Różne stężenia substancji wewnątrz i na zewnątrz komórki to jedna z najważniejszych oznak życia. Każda komórka, która jest w pełnej równowadze koncentracji ze swoim środowiskiem, jest martwa. To prawda, że wiele substancji, przede wszystkim woda, przenika do komórki zgodnie z gradientem ich stężeń. Istnieje jednak ogromna liczba substancji, które nie przestrzegają tej zasady.
Jeśli podzielimy różne substancje na klasy zgodnie z ich przepuszczalnością, to można wśród nich wyróżnić następujące główne grupy. Substancje niepolarne, charakteryzujący się równomiernym rozkładem elektronów między atomami tworzącymi cząsteczkę. Niekompletne połączenia biegunowe, w którym elektrony wiążące atomy są bardziej przyciągane do jednego z nich, w wyniku czego cząsteczka zamienia się w dipol. Związki jonowe, czyli elektrolity, gdzie elektrony walencyjne są związane tylko z jednym z atomów lub rodników. W ośrodkach o wysokiej stałej dielektrycznej wiązania te są znacznie osłabione, a poszczególne części cząsteczki zachowują się jak niezależne jony. Tutaj elektrolit dysocjuje na jony: dodatnie - aniony i ujemne - kationy.
Pomijając kwestię przenikania związków dwóch pierwszych klas do komórki, zajmijmy się bardziej szczegółowo naturą przenikania i rozmieszczenia różnych jonów między komórką a jej otoczeniem, ponieważ to właśnie ten rozkład leży u podstaw potencjał błonowy (MP) komórki.
Obecnie udowodniono eksperymentalnie, że zawartość ogniwa jest naładowana ujemnie w stosunku do jej powierzchni zewnętrznej i otoczenia. Ładunek ten nie jest taki sam w różnych ogniwach iw większości przypadków waha się od -50 do -100 mV (tabela 9). Obecność ładunku elektrycznego wskazuje na nierówny rozkład elektrolitów po obu stronach błony powierzchniowej komórki, co potocznie nazywa się asymetrią jonową.
Tabela 9. Wartości potencjałów błonowych (MP) różnych komórek pobudliwych (według Latmanizova, 1965)
Jak asymetria jonowa zachodzi po przeciwnych stronach błony komórkowej? Aby wyjaśnić tę kwestię, konieczne jest rozważenie, przynajmniej w ogólnej formie, zachowania elektrolitów podczas przechodzenia przez jakąkolwiek membranę półprzepuszczalną.
Wszystkie rozpuszczone substancje, w tym zjonizowane, przemieszczają się (w przypadku nierównomiernego rozmieszczenia w roztworze) zgodnie z gradientem stężeń z obszarów o wyższym stężeniu do obszarów o niższym stężeniu. Jeśli obszary roztworu o nierównych stężeniach są oddzielone jakąś membraną, wówczas dyfuzja substancji rozpuszczonych jest mniej lub bardziej spowolniona. Składniki jonowe rozpuszczonych elektrolitów - aniony i kationy - często mają różną przepuszczalność, a co za tym idzie różną ruchliwość. Wynika to z wielkości i konfiguracji jonów, a także wielkości ich otoczek hydratacyjnych. Na przykład kation Na + i anion Cl - powstający podczas dysocjacji soli kuchennej mają różną ruchliwość. Ruchliwość i przepuszczalność Na+ jest znacznie mniejsza niż Cl-. Odpowiada to ich uwodnionym średnicom 2,14 Å dla Cl - i 3,4 Å dla Na + . Przy przenikaniu dyfuzyjnym przez membranę z obszaru o wyższym stężeniu elektrolitu do obszaru o niższym stężeniu, jony Cl - przemieszczą się na drugą stronę membrany znacznie szybciej niż jony Na+, tak że jedna z jej powierzchni przez pewien czas będzie naładowany dodatnio, a drugi - ujemnie.
Jeśli membrana okaże się przepuszczalna tylko dla jonów Cl - i nieprzepuszczalna dla jonów Na+, to nie ma już chwilowej, ale stałej różnicy potencjałów między roztworami po obu stronach membrany. Ta potencjalna różnica nazywa się różnica potencjałów koncentracji i można je obliczyć za pomocą równania Nernsta:
![]()
gdzie E jest różnicą potencjałów w mV; R jest stałą gazową; T to temperatura bezwzględna; n - wartościowość; C 1 - wyższe, C 2 - niższe z dwóch stężeń; F to liczba Faradaya.
Prawidłowości te dotyczą przypadku, gdy roztwory znajdujące się po obu stronach błony półprzepuszczalnej zawierają te same jony, ale w różnych stężeniach.
Jednak, jak wiadomo, protoplazma komórki i płyn pozakomórkowy różnią się nie tylko stężeniem, ale także składem jonów, więc pojęcie potencjału stężenia nie wystarcza do wyjaśnienia zdarzeń zachodzących w rejonie komórki membrana.
Rozważmy uproszczony przypadek, bliższy rzeczywistym stosunkom elektrolitów w ogniwie. Załóżmy, że pewna membrana oddziela dwa różne elektrolity NaCl i KCl o tym samym stężeniu:
Załóżmy również, że błona ta jest przepuszczalna zarówno dla anionów Cl - jak i dla obu kationów, ale jej przepuszczalność dla K + jest znacznie większa niż dla Na + . Wówczas K+, jako bardziej ruchliwy jon, rozkłada się równomiernie po obu stronach membrany znacznie szybciej niż Na+ i przez cały czas, w którym stężenia Na+ pozostają wyrównane, jedna ze stron membrany, a mianowicie ta, na której NaCl był pierwotnie zlokalizowany, będzie naładowany dodatnio względem przeciwnej strony. Powstający w ten sposób potencjał nazywany jest potencjałem chemicznym. Gdyby membrana była całkowicie nieprzepuszczalna dla Na+, potencjał ten stałby się stały.
W przypadku komórkowego MP wszystkie procesy są znacznie bardziej złożone. Przede wszystkim skład elektrolitowy środowiska wewnątrzkomórkowego i zewnątrzkomórkowego jest dość złożony i oprócz KCl i NaCl zawiera szereg innych jonów, wśród których szczególnie ważną rolę odgrywają duże aniony organiczne, niezdolne do penetracji błony komórkowej w tworzeniu potencjału błonowego. Przybliżone stężenia różnych jonów w komórkach mięśniowych ssaków podano w tabeli. dziesięć.

Tabela 10. Przybliżone stężenia jonów i potencjały równowagi we włóknie mięśniowym ssaków i płynie pozakomórkowym (według J. W. Woodbury, 1963)
* (Obliczony na podstawie potencjału błonowego za pomocą równania Nernsta.)
Jak widać z tabeli, środowisko zewnątrzkomórkowe charakteryzuje się stosunkowo wysokim stężeniem jonów Na+ i Cl- oraz niskim stężeniem jonów K+. Natomiast środowisko wewnątrzkomórkowe zawiera niewielkie ilości Na + i Cl - , ale ma wysoką zawartość K + . Ponadto komórka zawiera ponad 150 mM wysokocząsteczkowych anionów organicznych A - na 1 litr wody wewnątrzkomórkowej, najwyraźniej pochodzenia białkowego. Dlatego przy konstruowaniu dalszego rozumowania należy uwzględnić ten dodatkowy czynnik.
Na ryc. 20 to bardzo uproszczony schemat procesów zachodzących na błonie komórkowej. Rozważając schemat zakłada się, że membrana jest nieprzepuszczalna nie tylko dla anionów wielkocząsteczkowych A - , ale także dla jonów Na +.

Ryż. 20. Powstawanie potencjału transbłonowego pod wpływem gradientów stężeń. (Woodbury, 1963). Wielkości symboli w prawej i lewej kolumnie wskazują na względne stężenie jonów w płynie zewnątrz- i wewnątrzkomórkowym. Przerywane strzałki i kółka - kierunek ruchu jonów, K + , A - , Na + i Cl - podczas przesuwania K + lub Cl - przez pory membrany. Inne wyjaśnienia w tekście
Niech w pewnym początkowym momencie po obu stronach membrany ośrodek z rozpuszczonymi w nim jonami będzie elektrycznie obojętny, tj. dodatnie i ujemne cząstki po obu stronach membrany równoważą się nawzajem. Prześledźmy zachowanie cząstek zdolnych do przenikania przez „pory membrany”. * Ze względu na wysokie wewnątrzkomórkowe stężenie jonów K+, jony te będą dyfundować przez błonę komórkową od wewnątrz na zewnątrz zgodnie z gradientem stężenia. Aniony o dużej masie cząsteczkowej równoważące ładunek elektryczny jonów K+ nie mogą im towarzyszyć, ponieważ membrana jest dla nich nieprzepuszczalna. Jony Na+ nie mogą ich również zastąpić w środowisku wewnątrzkomórkowym, gdyż zgodnie z założeniem nie przenikają przez błonę z zewnątrz do wewnątrz. W rezultacie wokół membrany następuje częściowe oddzielenie ładunku. Jej zewnętrzna strona, w wyniku pojawienia się nadmiaru potasu w ośrodku pozakomórkowym, zaczyna ładować się dodatnio, a wewnętrzna, w wyniku pojawienia się wysokocząsteczkowych jonów A nie zneutralizowanych przez potas, staje się naładowana ujemnie. W obszarze membrany powstaje pole elektryczne, które zaczyna zakłócać trwający proces, ponieważ dyfundujące jony K + mają ładunek. Jak wiadomo, ładunki elektryczne o tej samej nazwie odpychają się od siebie, więc ładunek dodatni powstający w środowisku zewnątrzkomórkowym zaczyna przeciwdziałać dalszemu przepływowi jonów K+ z wnętrza komórki na zewnątrz. Zatem dyfuzja K+ z ogniwa na zewnątrz powoduje pojawienie się sił elektrostatycznych, które uniemożliwiają proces pierwotnej dyfuzji, tj. powstaje ujemne sprzężenie zwrotne, które ogranicza wyjście K+ z ogniwa. Gdy dodatni potencjał środowiska zewnętrznego w stosunku do odpowiednio wzrastającego wewnątrzkomórkowego potencjału ujemnego osiągnie określoną wartość, ustala się dynamiczna równowaga między liczbą jonów K+ opuszczających komórkę a liczbą tych jonów wchodzących do komórki. Różnica potencjałów odpowiadająca temu momentowi po obu stronach membrany jest zwykle oznaczana jako potencjał równowagi dla danego jonu. Z toku rozumowania jasno wynika, że jego wartość zależy od stosunku stężeń wewnętrznych i zewnętrznych tego jonu i można ją obliczyć z równania Nernsta.
* (Wyrażenie „pory membranowe” nie powinno być rozumiane dosłownie, jako otwory o określonej wielkości. Nierówna przepuszczalność membrany dla różnych cząstek jest najwyraźniej związana ze złożonym oddziaływaniem między cząsteczkami membrany i tymi cząsteczkami. Niemniej jednak wielkość penetrujących cząstek, jak zobaczymy w dalszej części, odgrywa znaczącą rolę.)
Podobne rozumowanie można przeprowadzić w odniesieniu do jonu Cl -. Ten anion zaczyna wnikać w warunkowym momencie początkowym z zewnątrz do wewnątrz ze względu na jego stosunkowo wysokie stężenie w płynie pozakomórkowym. W takim przypadku wewnątrz ogniwa powstaje ładunek ujemny, a na zewnętrznej powierzchni membrany ładunek dodatni. Podobnie jak w przypadku K + , powstaje ujemne sprzężenie zwrotne, które ostatecznie ogranicza przepływ jonów Cl - do komórki po osiągnięciu potencjału równowagi między środowiskiem zewnętrznym i wewnętrznym.
Naturalnie, proces wyjścia K + z komórki i proces wejścia jonów Cl - do komórki są w interakcji, a poziom ustalonej różnicy potencjałów jest określony przez całość zachodzących zdarzeń.
Nietrudno zrozumieć niektóre mechanizmy powstawania MF w spoczynkowej żywej komórce na powyższym schemacie. * Obliczenia ilościowe przy użyciu równania Nernsta pokazują, że jony Cl - i K + są rozłożone po obu stronach membrany w przybliżeniu zgodnie z wielkością MP, tak że gradienty elektryczne i stężenia równoważą się.
* (Komórka, która nie zdaje sobie sprawy ze swojej specyficznej aktywności, jest umownie nazywana komórką spoczynkową. Jak widać z tego, co dalej, pojęcie odpoczynku jest abstrakcją fizjologiczną.)
Faktycznie opisany mechanizm występowania MF mógłby mieć miejsce, gdyby błona komórkowa była całkowicie nieprzepuszczalna dla jonów Na+. Jednak eleganckie i rygorystyczne badania Hodgkina i Keynesa (Keynes, 1954; Hodgkin a. Keynes, 1955) z użyciem radioaktywnego izotopu sodu Na24 wykazały, że błona komórkowa jest przepuszczalna dla Na+, chociaż jej przepuszczalność jest około 50 razy mniejsza niż przepuszczalność dla K + i Cl - . Na mięśniu brzuchatym łydki żaby wykazano eksperymentalnie (Keynes, 1954), że około 10-11 M Na+ przenika do włókna mięśniowego na sekundę przez 1 cm2 powierzchni komórki. Przy takim „dopływie” Na+ do komórki bez odpowiedniego „wycieku” z niego, jego wewnątrzkomórkowe stężenie powinno w ciągu godziny podwoić się. Jednak tak się nie dzieje, chociaż koncentracja i gradienty elektryczne „zmuszają” Na+ do wejścia do komórki. Dlatego muszą istnieć pewne specjalne mechanizmy, które zapewniają przejście jonów Na+ przez membranę wbrew gradientowi elektrochemicznemu, czyli od wewnątrz na zewnątrz. Mechanizmy te nazywane są aktywnym transferem. Wdrożenie tych ostatnich wymaga stałego wydatkowania energii wytwarzanej w wyniku komórkowych procesów metabolicznych.
Niestety nadal nie jest jasne, w jaki sposób energia metabolizmu komórkowego jest wykorzystywana podczas aktywnego transferu, jednak istnieją dość wyraźne dane doświadczalne dotyczące udziału adenozynotrójfosforanu i niektórych innych fosforanów w tym procesie (Caldwell a. Keynes, 1957; Caldwell, Hodgkin , Keynes a. Shaw, 1960a; Hogdkin, 1964).
Bezpośredni związek między aktywnym transferem a metabolizmem został po raz pierwszy wykazany przez Hodgkina i Keynesa (Hodgkin a. Keynes, 1955) w badaniach nad aksonem olbrzymiej mątwy (sepia). Metodologia badań opierała się na wykorzystaniu radioizotopu sodu – Na 24 i pozwoliła na ujawnienie szeregu interesujących wzorców aktywnego transferu. Przede wszystkim okazało się, że inhibitory metabolizmu (dinitrofenol, cyjanek, azydek), blokując pewne ogniwa w metabolicznych reakcjach oksydacyjnych, niemal całkowicie hamują proces usuwania Na+ z komórki*. Podobnie działa spadek temperatury leku, znacznie obniżając poziom procesów metabolicznych (ryc. 21).
* (Ostatnio wykazano również blokujący wpływ azydku na aktywny transfer jonów Na+ w neuronach ruchowych rdzenia kręgowego kotów (Ito i Oshima, 1964).)

Ryż. Rys. 21. Uwalnianie jonów Na + z olbrzymiego aksonu (mątwy) po zmianie składu roztworu do płukania przez dodanie dinitrofenolu (DNP) (A) i zmianę temperatury (B) (Hodgkin i Keynes, 1955). Na odciętej - czas w minutach; wzdłuż osi y - uwalnianie radioaktywnego Na 24 z komórki, mierzone w impulsach na minutę. Całkowita produkcja N + (radioaktywnego i konwencjonalnego) jest proporcjonalna do produkcji radioaktywnego Na +, z wyłączeniem tych zmian, które wynikają ze stopniowego rozcieńczania radioaktywnego Na + zwykłym w wyniku naturalnych procesów wymiany jonowej (ten stopniowy spadek stężenia radioaktywny Na + jest pokazany w kolumnie „woda morska”)
Ponadto stwierdzono, że wydalanie Na+ jest na ogół proporcjonalne do jego stężenia wewnętrznego i nie zależy od jego stężenia zewnętrznego. Jednocześnie wydajność Na + gwałtownie spada wraz ze spadkiem stężenia potasu w środowisku zewnętrznym i wzrasta wraz ze wzrostem zewnętrznego stężenia K + . Ta zależność jest również pokazana na ryc. 21. Dodatkowo okazało się, że trucizny metaboliczne również ograniczają przepływ K+ ze środowiska zewnętrznego do aksonu i że ten proces jest blokowany przez te wpływy niemal w takim samym stopniu jak usuwanie Na+ z komórki. Wszystko to pozwoliło stwierdzić, że mechanizm transportu aktywnego działa zgodnie z zasadą elektroneutralności, „wyprowadzając” jony Na+ z komórki i „wprowadzając” do komórki równoważną ilość jonów K+ (Hodgkin, 1958) . *
* (Później okazało się, że związek między wydalaniem Na + a wprowadzaniem K + do komórki podczas aktywnego transferu jest raczej swobodny, przynajmniej nie absolutnie sztywny (jeden do jednego), jak zakładano na początku (Caldwell, Hodgkin, Keynes a. Shaw, 1960a, 1960b). Do niedawna uważano, że transport aktywny jest determinowany przez poziom wewnętrznego stężenia Na+ (Hodkin i Keynes, 1956; Hodgkin, 1958). W ostatnich pracach Ecclesa i jego współpracowników dostarczono danych, że składnik potasowy aktywnego transportu jest aktywowany przez zmniejszenie wewnętrznego stężenia K i że jednocześnie KCl jest aktywnie transportowany do komórki (Eccles, 1964; Eccles, Eccles i Ito, 1964).)
Powstaje pytanie, w jaki sposób mechanizm oparty na zasadzie neutralności elektrycznej może powodować polaryzację membrany. Nie jest to trudne do zrozumienia, biorąc pod uwagę, że wewnątrz komórki zachodzi znaczna ilość wielkocząsteczkowych anionów A - i że przepuszczalność jonów Na + jest 50 razy mniejsza niż przepuszczalność jonów K +. Usuwany z komórki przez mechanizm aktywnego transferu sód powraca do komórki 50 razy wolniej na skutek dyfuzji niż potas wprowadzony do komórki opuszcza ją w postaci strumienia dyfuzyjnego. Ponadto potas dyfunduje z komórki wraz z neutralizującym ją jonem Cl -. W wyniku tych procesów zmniejsza się bezwzględna liczba jonów dodatnich wewnątrz komórki: Na + jest aktywnie wydalany i biernie towarzyszą jony Cl - potas. Nadmiar ładunków ujemnych powstaje w ogniwie z powodu obecności wysokocząsteczkowych jonów A -.
Jest to oczywiście bardzo uproszczony system rozumowania. Odzwierciedla jednak istotę zjawisk w sposób ogólny, nie dotykając niektórych komplikujących szczegółów, na przykład kwestii dystrybucji wody w tych warunkach.
Istnieje wiele różnych hipotez próbujących wyjaśnić mechanizm aktywnego transportu, ale żadna z nich nie została udowodniona doświadczalnie. Najbardziej atrakcyjne schematy z tzw. „przewoźnikiem pośrednim”. Przykładem jest schemat Shaw-Glynn (Glynn, 1957) (ryc. 22) Zaletą takiego schematu jest to, że nie implikuje on sztywnej zależności między wydalaniem Na+ a wejściem K+ do komórki. Tak więc zachowanie częściowego wydalania Na + przy całkowitym braku K + w środowisku zewnętrznym można wytłumaczyć faktem, że Na + ma również pewne powinowactwo do substancji X. Hipotetyczny schemat powinien wyjaśniać tę możliwość, ponieważ wydalanie Na+ jest rzeczywiście częściowo zachowane nawet przy całkowitym braku K+ w środowisku zewnątrzkomórkowym.

Ryż. 22. Hipotetyczny schemat „pompy” wymiany potasowo-sodowej. (Glynn, 1957). Zakłada się, że substancje X i Y działają wewnątrz błony. X ma duże powinowactwo do K + ; Y ma wysokie powinowactwo do Na + . X i Y poruszają się po błonie tylko w połączeniu z odpowiednim jonem
Do tej pory wszystkie zjawiska i mechanizmy związane z generowaniem MF, dla uproszczenia prezentacji, zostały opisane jedynie z jakościowego punktu widzenia. Jednak dla jaśniejszego wyobrażenia o zachodzących procesach wskazane jest podanie pewnych ilościowych charakterystyk polaryzacji membrany, zwłaszcza że pozwoli to również uzyskać jaśniejsze pojęcie o objętości zachodzącej wymiany jonowej procesy. *
* (Poniższy materiał cyfrowy pochodzi z Neurophysiology T. Ruh i wsp. (Ruch i wsp., 1963).)
Pojemność błony mięśnia poprzecznie prążkowanego żaby została określona doświadczalnie i wynosi około 10 mikrofaradów na cm2. Wielkość ładunku membrany - tego biologicznego kondensatora - jest określona przez wielkość różnicy potencjałów po obu stronach membrany (E m) i jej pojemność (C m): q = C m ⋅ E m. Bierzemy E m dla 90 mV. Następnie ładunek q \u003d 10 × 10 -6 f / cm 2 × 0,09 w \u003d 9 × 10 -7 kulombów na 1 cm 2. Jeśli przeliczymy uzyskaną wartość ładunku przez liczbę jonów biorących udział w jego tworzeniu, otrzymamy następujące wyniki.
Zgodnie z prawem Avogadro, 1 gramowa cząsteczka zawiera 6,023 × 10 23 cząsteczek. W tym przypadku mamy do czynienia z jonami jednowartościowymi zawierającymi 1 jednostkę. ładunek równy co do wielkości ładunkowi jednego elektronu. Ładunek elektronu wyrażony w kulombach wynosi 1,6 × 10 -19 . Wtedy ładunek jednego mola kationów jednowartościowych wynosi 1,6 × 10 -19 × 6,023 × 10 23 = 96500 kulombów. Ponieważ ładunek na błonie komórkowej włókna mięśniowego wynosi 9 × 10 -7 kulombów cm 2 , oznacza to, że błona ta oddziela tylko 9 × 10 -7 / 96500 = 9,5 × 10 -12 mol zjonizowanych jednowartościowych cząstek w 1 cm 2 sekcja. Jednocześnie 1 cm3 płynu wewnątrzkomórkowego zawiera 1,5 × 10 -6 mol kationów lub anionów, czyli kilka milionów razy więcej. Obliczenia pokazują, że warstwa płynu wewnątrzkomórkowego o grubości zaledwie 6 Å zapewnia wystarczającą ilość jonów do wytworzenia potencjału 90 mV przez błonę.
Wartości te wyjaśniają, jak niewielka ilość jonów z ich całkowitej liczby w komórce bierze udział w utrzymaniu MP i, jak zostanie pokazane poniżej, w generowaniu potencjału czynnościowego.
Należy również zauważyć, że komórka ma wystarczające zasoby energii do aktywnego transferu Na+. Keynes i Maisel (Keynes a. Maisel, 1954) wykazali eksperymentalnie, że zużycie energii na aktywny transfer Na+ w mięśniu szkieletowym żaby jest znacznie mniejsze niż całkowite zużycie energii komórki, obliczone na podstawie zużycia tlenu. * W większości eksperymentów z mięśniem w spoczynku, tylko około 10% wydatku energetycznego trafia do aktywnego wydalania Na+. Jeśli przyjmiemy, że sprawność aktywnego transferu wynosi tylko 50% lub nawet mniej, to zużycie energii na ten proces nadal stanowi stosunkowo niewielką część energii wytwarzanej przez ogniwo. Te dane ilościowe znacząco wspierają koncepcję mechanizmu aktywnego transferu, pokazując, że z punktu widzenia bilansu energetycznego komórki taki proces jest całkiem możliwy.
17 ..spoczynkowy potencjał błonowy (MPP) lub potencjał spoczynkowy (PP) nazywana jest różnicą potencjałów komórki spoczynkowej między wewnętrzną i zewnętrzną stroną błony.Wewnętrzna strona błony komórkowej jest naładowana ujemnie w stosunku do zewnętrznej. Przyjmując potencjał rozwiązania zewnętrznego jako zero, MPP jest rejestrowany ze znakiem minus. Wartość WFP zależy od rodzaju tkanki i waha się od -9 do -100 mV. Dlatego w spoczynku błona komórkowa spolaryzowane. Spadek wartości MPP nazywa się depolaryzacja zwiększać - hiperpolaryzacja, przywracanie pierwotnej wartości WFP-repolaryzacja membrany.
Główne postanowienia membranowej teorii pochodzenia WFP sprowadza się do następujących. W spoczynku błona komórkowa jest dobrze przepuszczalna dla jonów K + (w niektórych komórkach i dla SG), mniej przepuszczalna dla Na + i praktycznie nieprzepuszczalna dla białek wewnątrzkomórkowych i innych jonów organicznych. Jony K + dyfundują z komórki wzdłuż gradientu stężeń, podczas gdy niepenetrujące aniony pozostają w cytoplazmie, zapewniając pojawienie się różnicy potencjałów w poprzek błony.
Powstała różnica potencjałów zapobiega wychodzeniu K+ z komórki, a przy pewnej wartości następuje równowaga między wychodzeniem K+ wzdłuż gradientu stężenia a wejściem tych kationów wzdłuż wynikowego gradientu elektrycznego. Potencjał błonowy, przy którym osiąga się tę równowagę, nazywa się potencjał równowagi. Jego wartość można obliczyć z równania Nernsta:
10 We włóknach nerwowych sygnały są przekazywane przez potencjały czynnościowe, które są szybkimi zmianami potencjału błony, które szybko rozprzestrzeniają się wzdłuż błony włókna nerwowego. Każdy potencjał czynnościowy zaczyna się od szybkiego przesunięcia potencjału spoczynkowego z normalnej wartości ujemnej na wartość dodatnią, a następnie prawie równie szybko powraca do potencjału ujemnego. Kiedy sygnał nerwowy jest przewodzony, potencjał czynnościowy porusza się wzdłuż włókna nerwowego aż do końca. Rysunek przedstawia zmiany zachodzące na membranie podczas potencjału czynnościowego, z przeniesieniem ładunków dodatnich do włókna na początku i powrotem ładunków dodatnich na zewnątrz na końcu. Dolna część rysunku przedstawia graficznie kolejne zmiany potencjału błonowego w ciągu kilku 1/10000 s, ilustrując wybuchowy początek potencjału czynnościowego i prawie równie szybki powrót do zdrowia. etap odpoczynku. Ten etap jest reprezentowany przez spoczynkowy potencjał błonowy, który poprzedza potencjał czynnościowy. Membrana na tym etapie jest spolaryzowana z powodu obecności ujemnego potencjału błonowego wynoszącego -90 mV. faza depolaryzacji. W tym czasie membrana nagle staje się wysoce przepuszczalna dla jonów sodu, umożliwiając dyfuzję ogromnej liczby dodatnio naładowanych jonów sodu do aksonu. Normalny spolaryzowany stan -90 mV jest natychmiast neutralizowany przez nadchodzące dodatnio naładowane jony sodu, powodując gwałtowny wzrost potencjału w kierunku dodatnim. Proces ten nazywa się depolaryzacją.W dużych włóknach nerwowych znaczny nadmiar wewnętrznie dodatnich jonów sodu zwykle powoduje, że potencjał błonowy „przeskakuje” poza poziom zerowy, stając się lekko dodatnim. W niektórych mniejszych włóknach, jak w większości neuronów ośrodkowego układu nerwowego, potencjał osiąga poziom zerowy bez „przeskakiwania” go. faza repolaryzacji. W ciągu kilku ułamków milisekundy po gwałtownym wzroście przepuszczalności membrany dla jonów sodu, kanały sodowe zaczynają się zamykać, a kanały potasowe otwierać. W rezultacie szybka dyfuzja na zewnątrz jonów potasu przywraca normalny ujemny potencjał spoczynkowy błony. Proces ten nazywa się repolaryzacją błony. potencjał czynnościowy W celu pełniejszego zrozumienia czynników powodujących depolaryzację i repolaryzację konieczne jest zbadanie cech dwóch innych typów kanałów transportowych w błonie włókien nerwowych: elektrycznie sterowanych kanałów sodowych i potasowych. Sterowane elektrycznie kanały sodowe i potasowe. Niezbędnym uczestnikiem procesów depolaryzacji i repolaryzacji podczas rozwoju potencjału czynnościowego w błonie włókien nerwowych jest elektrycznie sterowany kanał sodowy. Elektrycznie bramkowany kanał potasowy również odgrywa ważną rolę w zwiększaniu szybkości repolaryzacji błony. Oprócz pompy Na+/K+ i kanałów przeciekowych K*/Na+ istnieją oba typy kanałów sterowanych elektrycznie. Kanał sodowy sterowany elektrycznie. W górnej części rysunku elektrycznie sterowany kanał sodowy pokazano w trzech różnych stanach. Ten kanał ma dwie bramki: jedną w pobliżu zewnętrznej części kanału, która nazywana jest bramką aktywacyjną, drugą - w pobliżu wnętrza kanału, która nazywana jest bramką inaktywacyjną. Górna lewa strona rysunku pokazuje stan spoczynkowy tej bramki, gdy spoczynkowy potencjał błonowy wynosi -90 mV. W tych warunkach bramki aktywacyjne są zamknięte i zapobiegają przedostawaniu się jonów sodu do włókna. aktywacja kanału sodowego. Gdy spoczynkowy potencjał błonowy przesuwa się w kierunku mniej ujemnych wartości, wzrastając od -90 mV do zera, na pewnym poziomie (zwykle między -70 a -50 mV) następuje nagła zmiana konformacyjna w bramce aktywacji, w wyniku , przechodzą w stan całkowicie otwarty. Ten stan nazywany jest stanem aktywnym kanału, w którym jony sodu mogą swobodnie wnikać przez niego do włókna; natomiast przepuszczalność membrany dla sodu wzrasta w zakresie od 500 do 5000 razy. Inaktywacja kanału sodowego. W prawym górnym rogu rysunku widać trzeci stan kanału sodowego. Wzrost potencjału, który otwiera bramkę aktywacyjną, zamyka bramkę dezaktywacyjną. Jednak bramka dezaktywacyjna zamyka się w ciągu kilku dziesiątych milisekundy po otwarciu bramki aktywacyjnej. Oznacza to, że zmiana konformacyjna prowadząca do zamknięcia bramki inaktywacji jest procesem wolniejszym niż zmiana konformacyjna, która otwiera bramkę aktywacji. W rezultacie kilka dziesiątych milisekund po otwarciu kanału sodowego bramka dezaktywacyjna zamyka się, a jony sodu nie mogą już wnikać do włókna. Od tego momentu potencjał błonowy zaczyna powracać do poziomu spoczynkowego, tj. rozpoczyna się proces repolaryzacji. Jest jeszcze inna ważna cecha procesu inaktywacji kanału sodowego: bramka inaktywacji nie otwiera się ponownie, dopóki potencjał błonowy nie powróci do wartości równej lub zbliżonej do poziomu początkowego potencjału spoczynkowego. W związku z tym ponowne otwarcie kanałów sodowych jest zwykle niemożliwe bez uprzedniej repolaryzacji włókna nerwowego.
13 Mechanizm przewodzenia wzbudzenia wzdłuż włókien nerwowych zależy od ich rodzaju. Istnieją dwa rodzaje włókien nerwowych: mielinowane i niezmielinizowane. Procesy metaboliczne we włóknach niezmielinizowanych nie zapewniają szybkiej kompensacji wydatku energetycznego. Rozprzestrzenianie się wzbudzenia będzie przebiegało ze stopniowym tłumieniem - ze spadkiem. Dekrementacyjne zachowanie pobudzenia jest charakterystyczne dla słabo zorganizowanego układu nerwowego. Wzbudzenie jest propagowane przez małe prądy kołowe, które występują wewnątrz włókna lub w otaczającej go cieczy. Pomiędzy obszarami wzbudzonymi i niewzbudzonymi powstaje potencjalna różnica, co przyczynia się do występowania prądów kołowych. Prąd rozprzestrzeni się od ładunku „+” do „-”. W punkcie wyjścia prądu kołowego zwiększa się przepuszczalność błony plazmatycznej dla jonów Na, co powoduje depolaryzację błony. Pomiędzy nowo wzbudzonym obszarem a sąsiednim obszarem niewzbudzonym ponownie powstaje różnica potencjałów, co prowadzi do występowania prądów kołowych. Wzbudzenie stopniowo obejmuje sąsiednie odcinki osiowego cylindra i w ten sposób rozprzestrzenia się na koniec aksonu. We włóknach mielinowych, dzięki doskonałości metabolizmu, pobudzenie przechodzi bez zaniku, bez ubytku. Ze względu na duży promień włókna nerwowego, ze względu na osłonkę mielinową, prąd elektryczny może wchodzić i opuszczać włókno tylko w obszarze przechwytywania. Po zastosowaniu podrażnienia następuje depolaryzacja w obszarze przecięcia A, sąsiedni punkt przecięcia B jest w tym momencie spolaryzowany. Pomiędzy przejęciami powstaje różnica potencjałów i pojawiają się prądy kołowe. Z powodu prądów kołowych wzbudzane są inne przechwyty, podczas gdy wzbudzenie rozprzestrzenia się w sposób słony, gwałtownie od jednego przejęcia do drugiego. Istnieją trzy prawa przewodzenia podrażnienia wzdłuż włókna nerwowego. Prawo integralności anatomicznej i fizjologicznej. Przewodzenie impulsów wzdłuż włókna nerwowego jest możliwe tylko wtedy, gdy jego integralność nie jest naruszona. Prawo izolowanego przewodzenia wzbudzenia. Istnieje szereg cech rozprzestrzeniania się pobudzenia we włóknach nerwów obwodowych, miazgi i pozapłucnych. We włóknach nerwów obwodowych pobudzenie jest przekazywane tylko wzdłuż włókna nerwowego, ale nie jest przekazywane do sąsiednich włókien nerwowych znajdujących się w tym samym pniu nerwu. We włóknach nerwowych miazgi rolę izolatora pełni osłonka mielinowa. Dzięki mielinie wzrasta rezystywność, a pojemność elektryczna powłoki maleje. W niemięsistych włóknach nerwowych pobudzenie jest przekazywane w izolacji. Prawo obustronnego wzbudzenia. Włókno nerwowe przewodzi impulsy nerwowe w dwóch kierunkach - dośrodkowo i odśrodkowo.
14 synapsy - Jest to wyspecjalizowana struktura, która zapewnia przekazywanie impulsu nerwowego z włókna nerwowego do komórki efektorowej - włókna mięśniowego, neuronu lub komórki wydzielniczej.
synapsy- są to połączenia wyrostka nerwowego (akson) jednego neuronu z ciałem lub wyrostkiem (dendryt, akson) innej komórki nerwowej (przerywany kontakt między komórkami nerwowymi).
Wszystkie struktury, które zapewniają transmisję sygnału z jednej struktury nerwowej do drugiej - synapsy .
Oznaczający- przekazuje impulsy nerwowe z jednego neuronu do drugiego => zapewnia transmisję wzbudzenia wzdłuż włókna nerwowego (propagacja sygnału).
Duża liczba synaps zapewnia duży obszar do przesyłania informacji.
Struktura synaps:
1. błona presynaptyczna- należy do neuronu, z którego przesyłany jest sygnał.
2. szczelina synaptyczna, wypełniony cieczą o wysokiej zawartości jonów Ca.
3. błona postsynaptyczna- należy do komórek, do których przesyłany jest sygnał.
Między neuronami zawsze jest luka wypełniona płynem śródmiąższowym.
W zależności od gęstości membran występują:
- symetryczny(o tej samej gęstości membrany)
- asymetryczny(gęstość jednej z membran jest większa)
błona presynaptyczna obejmuje przedłużenie aksonu neuronu transmisyjnego.
Rozbudowa - przycisk synaptyczny/płytka synaptyczna;.
Na tabliczce - pęcherzyki synaptyczne (pęcherzyki).
Po wewnętrznej stronie błony presynaptycznej białko/heksagonalna siatka(wymagane do uwolnienia mediatora), w którym znajduje się białko - neuron . Wypełnione pęcherzykami synaptycznymi, które zawierają mediator- specjalna substancja zaangażowana w transmisję sygnału.
Błona pęcherzyka zawiera - stenina (białko).
błona postsynaptyczna obejmuje komórkę efektorową. Zawiera cząsteczki białka, które są selektywnie wrażliwe na mediatora tej synapsy, co zapewnia interakcję.
Cząsteczki te są częścią kanałów błony postsynaptycznej + enzymów (wiele), które mogą zniszczyć połączenie mediatora z receptorami.
Receptory na błonie postsynaptycznej.
Błona postsynaptyczna zawiera receptory związane z mediatorem tej synapsy.
Między nimi jest szczelina zatrzaskowa . Jest wypełniony płynem międzykomórkowym zawierającym dużą ilość wapnia. Posiada szereg cech strukturalnych – zawiera cząsteczki białka, które są wrażliwe na mediator przenoszący sygnały.
15 Opóźnienie synaptyczne w przewodzeniu wzbudzenia
Aby wzbudzenie rozprzestrzeniło się wzdłuż łuku odruchowego, poświęca się pewien czas. Okres ten składa się z następujących okresów:
1. okres czasowo niezbędny do wzbudzenia receptorów (receptora) i do przewodzenia impulsów wzbudzających wzdłuż włókien doprowadzających do środka;
2. okres czasu niezbędny do rozprzestrzeniania się wzbudzenia przez ośrodki nerwowe;
3. czas wymagany do propagacji wzbudzenia wzdłuż włókien odprowadzających do ciała roboczego;
4. okres utajony ciała roboczego.
16 Hamowanie odgrywa ważną rolę w przetwarzaniu informacji wprowadzanych do OUN. Ta rola jest szczególnie wyraźna w hamowaniu presynaptycznym. Dokładniej reguluje proces wzbudzania, ponieważ pojedyncze włókna nerwowe mogą być blokowane przez to hamowanie. Do jednego neuronu pobudzającego mogą docierać setki i tysiące impulsów poprzez różne terminale. Jednocześnie liczba impulsów docierających do neuronu jest określana przez hamowanie presynaptyczne. Zahamowanie dróg bocznych zapewnia selekcję istotnych sygnałów z tła. Blokada inhibicji prowadzi do szerokiego napromieniowania pobudzenia i drgawek, na przykład, gdy inhibicja presynaptyczna jest wyłączana przez bikukulinę.
Połączenie materii organicznej i metali w żywych istotach, wykorzystanie żywej komórki jako przewodnika prądu elektrycznego nie może nie zaskoczyć. Im więcej dowiaduję się o swoim ciele, tym częściej zaskakuje jego proste działanie. Zginanie ramienia może być postrzegane nie jako zwykły, prawie niezauważalny proces, ale w wyniku skoordynowanej pracy mięśni i ścięgien, które otrzymały polecenie z mózgu, przekazywane przez włókna nerwowe za pomocą elektryczności. Patrząc na pasmo głębokiego opalizującego błękitu, który pojawia się na kawałku wypolerowanego labradorytu, który obracam w dłoniach, odbieram fale elektryczne emitowane przez receptory moich oczu. W receptorach informacja ta jest zakodowana w postaci serii impulsów elektrycznych. Jest przekazywany do mózgu, przetwarzany przez niego, a wynikiem tego wszystkiego jest postrzeganie tajemniczo pięknego kamienia, emocji, które w tym czasie powstają. Matryca w skrócie :)
Wraz z podziwem pojawia się również chęć dowiedzenia się, jak to wszystko się dzieje, chęć zagłębienia się w szczegóły każdego procesu.
Potencjał czynnościowy to impuls elektryczny, który dostarcza informacje z receptorów do mózgu i za pomocą którego mózg kontroluje ciało. Jego istnienie wymaga specjalnie ułożonych komórek, neuronów, których błony są oblepione z obu stron dodatnio i ujemnie naładowanymi jonami, które tylko czekają na sygnał, aby ten impuls elektryczny przenieść na całej długości komórki. Głównym miejscem istnienia potencjału czynnościowego jest akson neuronu. Dendryty niektórych typów neuronów są również zdolne do przewodzenia impulsów elektrycznych.

Wystarczy spojrzeć na rysunek po lewej, aby zobaczyć główną różnicę między potencjałem spoczynkowym a potencjałem czynnościowym: gdy membrana jest w spoczynku, na całej jej długości na zewnątrz występuje dodatni ładunek elektryczny, a ujemny W środku.

A potencjał czynnościowy przechodzący przez neuron z ciała komórki do końca aksonu jest zmianą zewnętrznych i wewnętrznych ładunków błony komórkowej na niewielkim obszarze neuronu: przez bardzo krótki okres czasu ładunki ujemne i dodatnie po przeciwnych stronach błony zamieniają się miejscami.
Pojawiają się pytania: co wyzwala potencjał działania? Jaka jest jej fizyczna natura, co dokładnie dzieje się w komórce podczas jej istnienia? Jak się rozprzestrzenia? Dlaczego zmierza tylko w jednym kierunku? W jaki sposób komórka przywraca swój początkowy potencjał spoczynkowy po przejściu przez nią fali pobudzenia?
Więc pierwsze pytanie brzmi: Jak generowany jest potencjał działania? Ponieważ transmisja sygnałów z neuronów źródłowych do neuronów odbiorczych, lub neuron postsynaptyczny (neuron postsynaptyczny), występuje w synapsach, tam należy szukać sygnału wyzwalającego potencjał czynnościowy. Substancje sygnalizacyjne docierają do neuronu przez synapsę, ale ich przybycie niekoniecznie oznacza wywołanie wzbudzenia neuronu. Gdyby neuron reagował na bardzo małe ilości substancji sygnalizacyjnych, byłoby to tak niewygodne, jak posiadanie superczułego przełącznika, który zadziała, jeśli przypadkowo na niego kichniesz.

Potrzeba wystarczająco silnej ekspozycji na pewne substancje sygnalizacyjne, aby wyzwolić potencjał czynnościowy. W rzeczywistych warunkach na ciele neuronu zwykle powstają potencjały postsynaptyczne - tymczasowe zmiany ładunków błony spowodowane przez substancje sygnałowe, które przeszły przez synapsę.
Potencjały postsynaptyczne bardzo różnią się od potencjałów czynnościowych w przyrodzie. Jeśli potencjał czynnościowy ma jedną ściśle określoną wartość, to wartość potencjałów postsynaptycznych zależy tylko od rodzaju i ilości substancji chemicznych, które je wywołały, przenoszonych przez synapsy. Podczas gdy potencjały czynnościowe generowane przez różne neurony są w przybliżeniu takie same, potencjały postsynaptyczne występujące w różnych synapsach wejściowych tego samego neuronu różnią się znacznie pod względem wielkości i czasu trwania. W jednej synapsie neuronu przychodzący impuls nerwowy może spowodować zmianę potencjału błonowego o 0,1 mV, aw innej o 20 mV. Ponadto istnieją zarówno sygnały wywołujące potencjał czynnościowy, jak i hamujące.
Ciało komórki służy jako rodzaj kalkulatora, najprostszego komputera analogowego, który sumuje wszystkie wchodzące do niego sygnały w postaci powstających potencjałów postsynaptycznych. Komputery analogowe są bardzo proste. Na przykład możesz zbudować komputer analogowy wody. Jeśli do słoika z jednej rury wleje się trzy kilogramy wody, a z drugiej pięć kilogramów wody do tego samego słoika, to po zważeniu słoika i stwierdzeniu, że waga pokazuje wagę równą ośmiu kilogramom, dochodzimy do wniosku, że 3 + 5 = 8. U zarania rozwoju technologii komputerowej stosowano zarówno komputery wodne, jak i inne komputery analogowe, ale najbardziej zaawansowana była oczywiście maszyna, która sumuje prądy elektryczne dokładnie tak, jak neurony. Operatorzy takich komputerów zajmowali się obliczeniami, wtykając wtyczki z jednego gniazdka do drugiego, łącząc ze sobą różne prądy.
Tak więc w neuronie zachodzi ta sama suma prądów elektrycznych. W trakcie tego podsumowania można przejść dalej próg wzbudzenia (próg wzbudzenia): zostanie osiągnięta pewna wartość potencjału błonowego, która wyzwala potencjał czynnościowy. Zwróć uwagę, że aby osiągnąć próg wzbudzenia, potencjał błony musi wzrosnąć. Jeśli spoczynkowy potencjał błonowy neuronu wynosi -70 mV, to jego próg wzbudzenia może wynosić -55 mV. Spadek potencjału błonowego np. do -90 mV prowadzi do tego, że do pokonania progu wzbudzenia potrzebny jest silniejszy efekt, ponieważ pozostaje on taki sam: -55 mV.

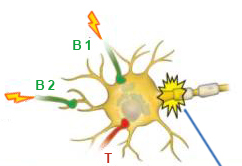
W neuronie postsynaptycznym, który odbiera sygnały z dwóch neuronów pobudzających i jednego hamującego, mogą występować różne kombinacje sygnałów i nie każdy z nich prowadzi do wystąpienia potencjału czynnościowego.
Na pierwszym schemacie powyższego rysunku przedstawiona jest sytuacja, w której słaby sygnał pochodzi tylko z jednego neuronu wzbudzającego, co nie wystarcza do przekroczenia progu wzbudzenia. Ten sygnał jest powtarzany, ale do czasu, gdy nadchodzi drugi, równie słaby sygnał z tego samego aksonu, wzbudzenie błony osiągnęło już potencjał spoczynkowy i nie powstaje żaden potencjał czynnościowy.
Drugi wykres pokazuje ten sam słaby sygnał, ale nadchodzący z wyższą częstotliwością. To właśnie ze względu na wzrost częstotliwości sygnału membrana nie ma czasu na powrót do stanu spoczynku w przerwie między dwoma sygnałami. W tym przypadku suma ładunku resztkowego błony po pierwszym sygnale i po drugim sygnale umożliwia pokonanie progu wzbudzenia neuronu. W tym przypadku u podstawy aksonu, który nazywa się wzgórek aksonów (akson pagórek ['aksɒn 'hɪlək]), pojawia się potencjał czynnościowy.
W trzecim schemacie słabe sygnały pochodzą z obu neuronów pobudzających, a ich suma wystarcza do wyzwolenia potencjału czynnościowego.
Czwarty schemat pokazuje wpływ sygnałów pobudzających i hamujących na neuron postsynaptyczny. Gdy odbierany jest tylko sygnał pobudzający, potencjał błonowy nieznacznie wzrasta, a gdy odbierany jest tylko sygnał hamujący, zmniejsza się. Jeśli te dwa sygnały nadejdą w tym samym czasie, to po prostu niszczą się nawzajem, pozostawiając potencjał błonowy w stanie zbliżonym do potencjału spoczynkowego. Przypomina to naciskanie na te same drzwi z różnych stron: osobno każde uderzenie mogłoby lekko otworzyć drzwi lub odwrotnie, zamknąć je, ale istniejące jednocześnie, prowadzą tylko do nieznacznego przesunięcia w kierunku działania większej siły .
Natura systemu jest taka, że nawet małe potencjały postsynaptyczne, zsumowane, mogą wywołać duży efekt. Neuron „sumuje” wszystkie pobudzające potencjały postsynaptyczne, które do niego weszły, „odejmuje” nadchodzące hamujące potencjały postsynaptyczne, a gdy tylko próg wzbudzenia zostanie przekroczony, powstaje potencjał czynnościowy.
Aby zrozumieć, co dzieje się w komórce podczas wyzwalanie i przekazywanie potencjału czynnościowego, należy wymienić każdego uczestniczącego gracza i jego działanie, tak jak to zrobiono przy opisie występowania spoczynkowego potencjału błonowego w . Więc, aktorów w tworzeniu potencjału działania:

1 . , które były zamknięte, gdy istniał spoczynkowy potencjał błonowy, otwierały się, gdy tylko potencjał osiągnął próg wzbudzenia. Ponieważ na membranie wewnątrz ogniwa znajduje się ładunek ujemny, w tej chwili
2 . pozytywnie naładowany jony sodu (Na+). To oni wytworzyli ładunek dodatni na zewnętrznej powierzchni błony neuronu i w tym momencie przenoszą go do środka, pozostawiając na błonie zewnętrznej ładunek ujemny, który tworzą ujemnie naładowane jony chlorkowe znajdujące się na zewnątrz komórki .
3 . w ten sposób jony chlorkowe (Cl -) tworzą ładunek ujemny na zewnętrznej powierzchni membrany.

Przemieszczanie się jonów sodu do komórki prowadzi do gwałtownej zmiany ładunków wewnątrz i na zewnątrz neuronu. Miejsce ujemnego ładunku wewnętrznego błony, który istnieje w pozostałej części neuronu, zajmuje dodatni ładunek jonów sodu. A poza błoną komórkową, po odejściu Na+ stamtąd, może w końcu zamanifestować się Cl -, którego ujemny ładunek nie jest już kompensowany przez dodatnio naładowane jony. W ten sposób potencjał błony komórkowej nagle wzrasta z -70 do +40 mV. To jest ten wzrost, zwany depolaryzacja (depolaryzacja) jest przedstawiony przez ostro wznoszącą się część szczytu na tym zdjęciu. Ten wzrost potencjału błonowego jest sygnałem do zamknięcia kanałów sodowych bramkowanych napięciem, a także otwarcia kanałów potasowych bramkowanych napięciem, o czym wcześniej nie mówiliśmy.
4 . kanały potasowe bramkowane napięciem . Występują one w neuronie oprócz kanałów przepływu potasu, które, jak zapewne pamiętasz z ostatniego rozdziału o potencjale, nie posiadają bramek i zapewniają stały odpływ potasu z komórki, odgrywając zasadniczą rolę w tworzeniu potencjał spoczynkowy. Okazuje się więc, że istnieją dwa rodzaje kanałów potasowych, a teraz przyjrzymy się działaniu kanału potasowego bramkowanego napięciem. Otwierające się po osiągnięciu szczytu potencjału czynnościowego, bramkowane napięciem kanały potasowe dramatycznie zwiększają wypływ jonów potasu z komórki. Dodatni ładunek, który istnieje w tym momencie wewnątrz neuronu, powstały w wyniku napływu Na +, aktywnie wypycha dodatnio naładowane jony potasu (K +) z komórki przez oba typy kanałów.

Prześledźmy więc jeszcze raz łańcuch zdarzeń aż do tego momentu: potencjał osiąga próg wzbudzenia, a ten sygnał elektryczny prowadzi do tego, że kanały sodowe zależne od napięcia otwierają się, jony Na + pędzą do wewnątrz, przyciągane przez ujemnie naładowane białek, następuje depolaryzacja, czyli różnica ładunków na zewnątrz i wewnątrz gwałtownie spadła, a nawet więcej - brak dodatnio naładowanych jonów sodu doprowadził do pojawienia się ładunku ujemnego tam, gdzie akurat był dodatni - nastąpiła zmiana biegunów. Działanie impulsu elektrycznego zakończyło się, kanały sodowe ponownie się zamknęły, a stłoczone wewnątrz jony Na+ wypchnęły podobnie naładowane jony K+ przez otwarte, zależne od napięcia kanały potasowe i przez kanały przepływu potasu.
Ładunek wewnętrznej powierzchni błony komórkowej zaczyna gwałtownie spadać, co na wykresie odpowiada dolnej części piku. Ten spadek opłat nazywa się repolaryzacja (repolaryzacja), czyli różnica w ładunkach na zewnątrz i wewnątrz neuronu zostaje przywrócona, tak jak początkowo była w spoczynku. Ale to różnica ładunku została przywrócona, a nie skład jonowy, który ją stworzył. Pomimo faktu, że pod koniec repolaryzacji na zewnątrz membrany powstaje ładunek dodatni, równy dodatniemu ładunkowi membrany w spoczynku, membrana jest nadal daleka od osiągnięcia stanu spoczynku. W końcu ten dodatni ładunek został tam wniesiony przez jony K +, które opuściły komórkę, a w stanie spoczynku podtrzymują go jony Na +, które obecnie są zamknięte w komórce, dokładnie w miejscu, w którym wcześniej znajdowały się jony potasu.
Pod koniec repolaryzacji może wystąpić spadek potencjału błonowego nawet poniżej jego wartości dla neuronu w spoczynku lub hiperpolaryzacja (hiperpolaryzacja [ˌhaɪpəpələraɪ'zeɪʃn]). Po krótkim czasie kompensuje się to dodatkowym uwalnianiem jonów potasu z komórki i następuje wcześniejszy, stabilny stan spoczynku.

Gdy potencjał błonowy staje się równy potencjałowi spoczynkowemu (-70 mV), kanały potasowe bramkowane napięciem zamykają się. Przez płynące kanały potasowe K+ nie wypływa już tak szybko, bo jego stężenie wewnątrz komórki spadło, a na zewnątrz znajduje się wiele jonów potasu. Jednocześnie w środku znajduje się wiele jonów sodu, które muszą zostać usunięte z komórki, aby potencjał czynnościowy mógł przez nią ponownie przejść.
Za przywrócenie pozycji wyjściowej „sód na zewnątrz, potas w środku” odpowiada pompa sodowo-potasowa, która usuwa z komórki trzy Na+ i jednocześnie wprowadza dwa K+ do środka. Jego praca została opisana w poprzednim rozdziale o potencjale , i jest to pokazane schematycznie w czwartej części tego rysunku. Połączone działanie pompy sodowo-potasowej i kanałów przepływu potasu prowadzi do tego, że po zewnętrznej stronie błony ponownie pojawia się duże stężenie Na+, a stężenie K+ wewnątrz neuronu jest wyższe niż na zewnątrz .
Gdyby działały tylko kanały przepływu potasu, to jony potasu tylko opuściłyby komórkę i niemożliwe byłoby utrzymanie większego stężenia K+ wewnątrz niż na zewnątrz. Gdyby tylko kanały sodowe zależne od napięcia były aktywne, jony sodu nie mogłyby w ogóle opuścić komórki po wejściu do niej, tworząc potencjał czynnościowy. Ale pompa sodowo-potasowa (to znaczy kanał sodowo-potasowy) rozwiązuje oba te problemy: wydobywa Na +, przywracając jego wysokie stężenie z zewnątrz. I wnika do wnętrza K+, który przeszedł przez kanały przepływowe, uniemożliwiając ich równomierne rozłożenie po obu stronach membrany, tracąc w ten sposób ciśnienie cząstkowe niezbędne do wytworzenia spoczynkowego potencjału membrany.
Jedna tysięczna sekundy po osiągnięciu minimum repolaryzacji błony neuronalnej aktywna praca kanałów błony przywraca potencjał spoczynkowy. Neuron jest gotowy do przewodzenia kolejnego potencjału czynnościowego.
5 . pozytywnie naładowany jony wapnia (Ca 2+) odgrywają ważną rolę w tworzeniu potencjału czynnościowego tylko w niektórych komórkach nerwowych, dlatego w tym rozdziale, który zawiera informacje wspólne dla różnych neuronów, nie zostanie on opisany.

Na tym kończy się ogólny opis ruchu jonów przez błonę komórkową, który przekształca potencjał spoczynkowy neuronu w potencjał czynnościowy. Można zrobić sobie przerwę i strawić to wraz z czymś smacznym, jednocześnie odżywiając mózg, rekompensując kalorie, które poświęcił na przetwarzanie tych wszystkich informacji.
Propagacja potencjału czynnościowego na całej długości aksonu wynika to z faktu, że prąd elektryczny, który powstał podczas jego istnienia w jednej sekcji błony neuronowej, pobudza sąsiednie sekcje, otwierając kanały zależne od napięcia sodu i wyzwalając już tam potencjał czynnościowy. Dokładniej, prąd elektryczny wyzwala potencjał czynnościowy tylko po jednej stronie miejsca, w którym istniał poprzedni potencjał. Ponieważ z drugiej strony kanały sodowe ulegają inaktywacji po niedawnym odkryciu, a bez ich udziału wystąpienie potencjału czynnościowego jest niemożliwe. Gdyby nie ten czas przerwy niefunkcjonalności pomiędzy dwoma otworami kanału sodowego, potencjał czynnościowy nie mógłby się rozprzestrzeniać tylko w jednym kierunku. Powstała reakcja łańcuchowa otwarcia kanału sodowego zapewnia przekazanie impulsu nerwowego z jego źródła do zakończeń aksonów odpowiednich dla innych neuronów (lub komórki mięśniowej).

Schematycznie tę reakcję łańcuchową można przedstawić w następujący sposób:
Czas 1: Prąd elektryczny, który powstał w wyniku otwarcia zależnych od napięcia kanałów sodowych w sekcji 1 wzbudza sąsiednie sekcje membrany.
Punkt w czasie 2: W wyniku tego wzbudzenia w miejscu 2 otwierają się zależne od napięcia kanały sodowe i powstaje potencjał czynnościowy, który z kolei wzbudza sąsiednie odcinki membrany. W miejscu 1 otworzyły się zależne od napięcia kanały potasowe, przywracając potencjał błonowy po przejściu potencjału czynnościowego.
Punkt w czasie 3: Prąd elektryczny wynikający z otwarcia kanałów sodowych bramkowanych napięciem w miejscu 2 spowodował otwarcie tych samych kanałów w miejscu 3 i tam przeniósł się potencjał czynnościowy. W miejscu 1 kanały sodowe bramkowane napięciem nie otworzyły się pomimo stymulacji elektrycznej z miejsca 2, ponieważ zostały tymczasowo dezaktywowane po wygenerowaniu potencjału czynnościowego. W miejscu 2 otworzyły się kanały potasowe bramkowane napięciem, przywracając spoczynkowy potencjał błonowy po przejściu potencjału czynnościowego.
Wiemy już, że wyrostki neuronów mogą być pokryte owiniętymi wokół nich komórkami neurogleju lub otoczką mielinową. To ona sprawia, że akson wygląda jak wydłużony łańcuch kiełbasek, w którym omówiono jego powstawanie i funkcje. Przekazywanie potencjału wzbudzenia wzdłuż "nagich", niepokrytych mielinowanymi komórkami włókien nerwowych różni się od ich przejścia wzdłuż zmielinizowanego aksonu. W porównaniu można zauważyć znaczące korzyści, jakie dają komórki neurogleju owinięte wokół aksonu:

1 . Rentowność. Błona aksonu pod mieliną nie ma kanałów do przejścia jonów, które tworzą potencjał czynnościowy. Kanały istnieją tylko w wąskich przestrzeniach między komórkami glejowymi, które nazywane są przechwyty Ranvier(zwężenie Ranviera) i tylko w tych wąskich odcinkach aksonu powstaje potencjał czynnościowy. Stężenie kanałów jonowych w tych przecięciach jest 100 razy większe niż w błonach włókien niezmielinizowanych, jednak do zapewnienia ich działania potrzeba znacznie mniej energii w porównaniu do tego, ile energii potrzeba by zapewnić kanały jonowe zlokalizowane na całej powierzchni powierzchnia nagi akson. Ponadto ta ekonomia umożliwia aksonowi szybsze „ładowanie” do następnego potencjału czynnościowego, dzięki czemu zmielinizowane włókna są w stanie przekazywać sygnały o wyższej częstotliwości.
2
. Prędkość. Na „gołym” aksonie sygnał jest przesyłany w każdym z jego punktów. Kanały jonowe otwierają się w każdym punkcie jego powierzchni, jeden po drugim, co przypomina uważne, skrupulatne wczytanie się w tekst.
W zmielinizowanym aksonie potencjał czynnościowy, który powstał w jednym węźle Ranviera, dociera do sąsiednich węzłów swoim polem elektrycznym, co prowadzi do pojawienia się w nich nowych potencjałów czynnościowych. Oznacza to, że pobudzenie przechodzi nagle od jednego przechwycenia do drugiego. W przypadku uszkodzenia jednego przechwycenia Ranviera, potencjał czynnościowy wzbudza kolejne przechwycenie po uszkodzonym, a jeśli to konieczne, nawet trzecie, czwarte i piąte z rzędu. Rozprzestrzenianie się pola elektrycznego na tej długości jest możliwe dzięki izolacji elektrycznej wytworzonej przez osłonki mielinowe: zmniejsza to rozpraszanie pola elektrycznego, które jest nieuniknione dla włókien niemielinowych.
Zatem szybkość propagacji potencjału czynnościowego wzdłuż włókien mielinowych jest znacznie wyższa w porównaniu z włóknami niezmielinizowanymi. Wszakże w tym samym czasie potencjał czynnościowy na „gołym” aksonie wzbudzi tylko obszar znajdujący się bezpośrednio obok niego, a potencjał czynnościowy na zmielinizowanym włóknie przeskoczy na odległość równą odległości między jednym lub więcej węzłów Ranviera:


Zakres szybkości transmisji potencjału czynnościowego w różnych zmielinizowanych włóknach jest bardzo szeroki: od kilku metrów na sekundę do „rekordu świata” ustanowionego przez akson krewetki: przewodzi wzbudzenie z prędkością przekraczającą 200 m/s.
Aby wyobrazić sobie, jak skutecznie można zwiększyć szybkość przewodzenia potencjału czynnościowego dzięki osłonce mielinowej, wystarczy porównać szybkość jej propagacji wzdłuż niezmielinizowanych i zmielinizowanych włókien ludzkich neuronów. Jeżeli przyjmiemy prędkość przewodzenia impulsów w równie cienkich włóknach, to we włóknie niezmielinizowanym będzie to około 1 m/s, a we włóknie zmielinizowanym 15-20 m/s. W grubych aksonach ludzkich pokrytych osłonką mielinową prędkość przewodzenia może sięgać 120 m/s.
Więc podsumujmy to: przekazywanie impulsu elektrycznego przez ciało i procesy neuronu odbywa się poprzez przepuszczanie jonów przez błonę komórkową.

1 . najpierw w błonie komórkowej powstaje potencjał spoczynkowy. Powstaje dodatnio naładowane jony sodu ( Na+) poza celą i białka naładowane ujemnie wewnątrz komórki. Cząstki tworzące ładunki znajdują się bardzo blisko błony komórkowej, ponieważ mają różne znaki i są do siebie przyciągane. Dlatego potencjał spoczynkowy istnieje tylko w pobliżu błony, ogólnie środowisko wewnątrz i na zewnątrz neuronu jest elektrycznie obojętne. Oprócz Na+ i ujemnie naładowanych białek biorą udział w tworzeniu i utrzymywaniu ładunku elektrycznego. dodatnio naładowane jony ( K + ) oraz ujemnie naładowane jony chlorkowe ( Cl-) również blisko membrany. Potencjał spoczynkowy wynosi około -70 mV (poniżej, przy wskazywaniu wartości liczbowej potencjałów błonowych, słowo „w przybliżeniu” zostanie pominięte).
2 . gdy neuron odbiera sygnały od innych neuronów poprzez synapsy pod ich wpływem na błonę ciała neuronu, potencjały postsynaptyczne, czyli zmiany wielkości ładunków elektrycznych na wewnętrznej i zewnętrznej powierzchni membrany. Pod wpływem tych sygnałów wewnętrzny ładunek ujemny i zewnętrzny ładunek dodatni mogą się zwiększać lub zmniejszać o różne wartości. A neuron działa jak kalkulator podsumowujący te efekty.
3 . wynikiem sumowania potencjałów postsynaptycznych może być kontynuacja neuronu w stanie spoczynku. Ale jeśli suma osiągnie wartość zwaną próg wzbudzenia, następnie w wzgórku aksonu (pogrubienie ciała neuronu u podstawy aksonu) potencjał czynnościowy.
4 . odgrywają ważną rolę w tworzeniu potencjału czynnościowego Błona komórkowa. Reguluje przechodzenie przez nią jonów (wszystkie powyższe, z wyjątkiem białek naładowanych ujemnie, które nie mogą opuścić komórki), otwierając i zamykając wyspecjalizowane kanały, które mogą przewodzić tylko określone jony.
5 . gdy błona neuronu przekracza próg wzbudzenia, otwiera się kanały sodowe bramkowane napięciem. Przeszkoda, która oddzielała dodatnio naładowany Na + od ujemnie naładowanych białek, które je przyciągały, znika i Na + pęd do celi. W ten sposób zewnętrzny ładunek dodatni jest przenoszony do wnętrza ogniwa, a ładunek ujemny wytworzony przez jony Cl - pozostaje na zewnątrz. Następuje zmiana potencjału błonowego z potencjału spoczynkowego od -70 mV do +40 mV i nazywa się to potencjał czynnościowy. Ważne jest, aby zrozumieć, że ta zmiana potencjału występuje tylko w obszarze membrany, w którym otworzyły się kanały sodowe bramkowane napięciem.
6 . W chwili, gdy błona osiąga potencjał 40 mV, kanały sodowe bramkowane napięciem zamykają się i otwierają w tym samym obszarze. kanały potasowe bramkowane napięciem. Na zewnątrz w tym momencie znajduje się ładunek ujemny, a wewnątrz - dodatni, stworzony przez jony Na +, które tam weszły i jony K + już tam w dużych ilościach. Jony potasu są wypychane do środowiska otaczającego neuron przez otwarte kanały, co znacznie zmniejsza ładunek wewnętrzny błony i zwiększa ładunek zewnętrzny. Po obu stronach błony powstają ładunki o tej samej wielkości, które normalnie zachowałyby potencjał spoczynkowy. Ale ten stan komórki różni się od potencjału spoczynkowego tym, że jony potasu i sodu nie znajdują się na swoich miejscach: Na + jest wewnątrz, a K + na zewnątrz. Ponownie wszystko opisane dotyczy tylko jednego miejsca na błonie, w którym potencjał czynnościowy właśnie przestał istnieć.
7 . Do przemieszczenia dodatnio naładowanych jonów Na+ i K+ po bokach membrany potrzebnych do utrzymania potencjału spoczynkowego, wykorzystuje się pompa sodowo-potasowa. A to działanie wpływa również na ograniczony obszar błony: ten, w którym właśnie zamknęły się kanały potasowe bramkowane napięciem.
Jeden cykl pompy sodowo-potasowej polega na tym, że: wychwytuje trzy jony Na + z wnętrza komórki, przenosi je na zewnątrz, gdzie wychwytuje dwa jony K + i przenosi je do środka. Oznacza to, że kanał wymienia trzy jony Na + na dwa jony K +, a Na + może poruszać się tylko na zewnątrz, a K + tylko wewnątrz komórki. W ten sposób przywracane jest początkowe stężenie sodu na zewnątrz membrany i potasu wewnątrz. Pompa sodowo-potasowa usuwa z ogniwa trzy dodatnio naładowane jony, ale tylko dwa do niego wchodzą. W konsekwencji, z każdym jej cyklem, wewnętrzny ładunek dodatni membrany maleje, a zewnętrzny wzrasta. Ponadto wypływające jony potasu odprowadzają ładunki dodatnie z wewnątrz na zewnątrz. Powstaje pytanie: skoro potencjał błonowy był już równy potencjałowi spoczynkowemu po wejściu jonów potasu do komórki, to po co obniżać go jeszcze bardziej? Odpowiedź jest taka, że oprócz wspomnianych tutaj jonów istnieją inne, których udział w tworzeniu potencjału spoczynkowego jest mniej znaczący, a dla zrekompensowania ich wpływu konieczne jest takie wyrównanie potencjału błonowego. Wynikiem tego procesu jest osiągnięcie spoczynkowego potencjału membrany: ładunek strony wewnętrznej membrany jest ujemny, strona zewnętrzna jest dodatnia, różnica potencjałów wynosi -70 mV.
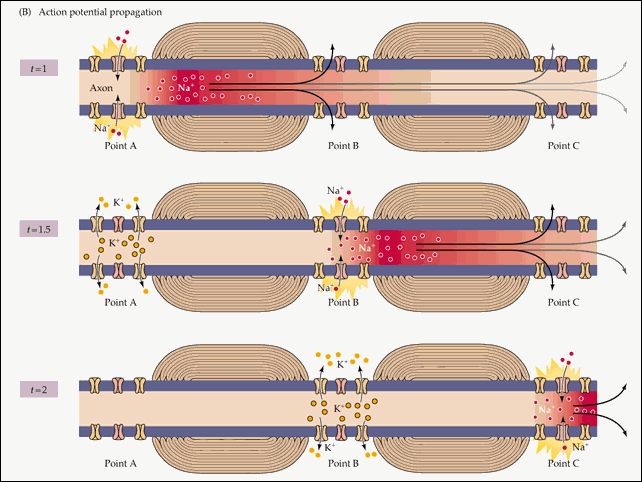
8 . Tak więc w tym miejscu błony komórkowej, gdzie zaledwie trzy kroki temu był potencjał czynnościowy, ponownie panuje potencjał spoczynkowy. Ale potencjał czynnościowy nie przeszedł bez śladu: jego istnienie (opisane w paragrafie 5) powstało pole elektryczne, które wzbudza sąsiadujące z nim części membrany. Z jednej strony (w kierunku końca aksonu) to wzbudzenie spowodowało otwarcie zależnych od napięcia kanałów sodowych, wyzwalając potencjał czynnościowy już w nowym obszarze. Co z kolei generuje pole elektryczne, które wyzwala kolejny potencjał czynnościowy, a cykl ten powtarza się na całej długości aksonu, od jego podstawy do końców połączonych z kolejnymi komórkami w łańcuchu nerwowym.
Po drugiej stronie potencjału czynnościowego, bliżej ciała neuronu, pomimo oddziaływania pola elektrycznego, potencjał czynnościowy nie powstaje. Ponieważ kanały sodowe bramkowane napięciem w tym obszarze zostały niedawno otwarte i na pewien czas są dezaktywowane. To właśnie ta przerwa w ich pracy sprawia, że potencjał czynnościowy rozchodzi się tylko w jednym kierunku.
9 . Każdy sygnał, niezależnie od tego, czy jest to transmisja informacji z neuronu czuciowego do mózgu, czy polecenie mózgu kierowane do neuronu ruchowego, jest przesyłany potencjały czynnościowe tej samej wielkości. Wynik potencjału czynnościowego (pojawienie się percepcji zapachu w wyniku pracy neuronów czuciowych lub ruch ciała bliżej jego źródła w wyniku skoordynowanej pracy neuronów ruchowych) zależy wyłącznie od tego, w którym obwodzie nerwowym znajduje się neuron. zawarte w tym potencjalnym przekazywaniu. Aby przekazać intensywność sygnału (siła zapachu lub wielkość skurczu mięśni), wykorzystywana jest częstotliwość, z jaką potencjały czynnościowe są przesyłane przez neuron. Słaby sygnał jest przesyłany przez rzadkie, a silny sygnał przez częste potencjały czynnościowe..
10 . otoczka mielinowa wokół procesów neuronu zwiększa tempo propagacji potencjału czynnościowego ze względu na to, że sygnał jest przesyłany na dużą odległość, równą szerokości jednej komórki neurogleju owiniętej wokół procesu, a nie do obszaru bezpośrednio sąsiadującego z obszarem wzbudzonym, jak to ma miejsce we włóknach niezmielinizowanych.
Pochodzenie mózgu Saveliev Sergey Vyacheslavovich
§ 8. Ładunki błony komórek nerwowych
Jednak główne właściwości układu nerwowego wynikają z umiejętności szybkiego reagowania na zmianę sytuacji wewnątrz lub na zewnątrz ciała. Procesów szybkich nie da się przeprowadzić zgodnie z powolnymi prawami humoralnymi, zachodzą one zgodnie z prawami elektrochemicznymi. Komórki nerwowe są w stanie odbierać, przechowywać, przetwarzać i przekazywać informacje za pomocą specjalnej aktywności elektrycznej. Posiadają ładunek błonowy - potencjał spoczynkowy i mogą go zmienić w potencjał czynnościowy, który rozprzestrzenia się z dużą prędkością w całym ciele komórki.
Potencjał spoczynkowy komórek nerwowych opiera się na równowadze sił elektrochemicznych i osmotycznych, które działają na granicy komórki - błonie. Błona komórkowa jest półprzepuszczalna. Oznacza to, że nie wszystkie substancje mogą przez nią przeniknąć. Membrana jest zawsze przepuszczalna dla wody, przepuszczalna selektywnie dla niektórych jonów i nieprzepuszczalna dla większości związków organicznych. Cząsteczki DNA, RNA, białek i aminokwasów znajdują się wewnątrz komórki i nie mogą swobodnie dyfundować przez błonę. Zgodnie z prawami osmozy woda musi dostać się do komórki. Ponieważ błona komórkowa jest nieprzepuszczalna dla cząsteczek organicznych, nie można osiągnąć równowagi osmotycznej. Komórka powinna pęknąć. Tak się nie dzieje, ponieważ siłom osmotycznym nieustannie przeciwstawiają się siły o zupełnie innym charakterze.
Siły te nie są osmotyczne, ale elektrochemiczne. Praca sił osmotycznych jest równoważona pracą sił elektrochemicznych. Z jednej strony nie pozwala to na pęknięcie komórki, a z drugiej jest źródłem stałego ładunku błony komórek nerwowych. Wewnątrz komórki znajdują się cząsteczki DNA, RNA, białek, aminokwasów i węglowodanów, które mają stały ładunek. Z reguły ładunek ten jest ujemny, a cząsteczki organiczne są zbiorem anionów wewnątrzkomórkowych (A). Ich ładunek jest równoważony wewnątrz ogniwa przez dodatnio naładowane jony potasu (K+). Na zewnątrz ogniwa głównym anionem jest chlor (Cl-), a kationem jest sód (Na+). W abstrakcyjnej sytuacji idealnej stężenia jonów wyrównałyby się w wyniku dyfuzji przez membranę. Jednak aniony wewnątrzkomórkowe są nieruchome, a specjalne kanały dla wszystkich jonów ruchomych są zwykle zamknięte. Co więcej, wyspecjalizowane kanały jonowe stale wypompowują nadmiar sodu i chlorku z komórki i pompują pozakomórkowy potas do komórki. Proces ten odbywa się z nakładem energii. Poświęca się na wytworzenie takiej wartości ładunku błonowego, że wystarczy przeciwdziałać siłom osmotycznym, które dążą do zniszczenia komórki.
W prawdziwej komórce główne problemy z siłami osmotycznymi (hydrostatycznymi) wynikają z różnic w stężeniu jonów potasu i sodu po obu stronach błony. jakąś rolę w tym
Proces ten odgrywają jony sodu i ruchliwość wody swobodnie przemieszczającej się przez błonę komórkową. Niemniej jednak potas ma podstawowe znaczenie, ponieważ różnice w jego stężeniu są maksymalne. Wewnątrz komórki jest około 40 razy więcej potasu i 9 razy mniej sodu niż w przestrzeni międzykomórkowej, więc potas ma tendencję do równoważenia sytuacji, przesuwając się wzdłuż gradientu stężenia z komórki, a sodu do komórki. Ponieważ znane są stężenia tych jonów wewnątrz i na zewnątrz komórki, procesy te można wyrazić w rzeczywistych wielkościach fizycznych. Praca, którą należy wykonać, aby zapobiec przemieszczaniu się jonów potasu poza komórkę wzdłuż gradientu stężenia (A 0), będzie równa:
gdzie R- stała gazowa; T- temperatura absolutna; [K + ] - stężenie molowe potasu w środku (w) i poza komórką (na zewnątrz).
Tylko siły elektryczne mogą przeciwdziałać siłom osmotycznym, które dążą do usunięcia potasu z komórki. Równowagę można osiągnąć tylko wtedy, gdy praca sił osmotycznych i elektrycznych jest równa. (Ae):
Dla każdego jonu można obliczyć pracę potrzebną do pokonania równowagi sił elektrycznych powstających w wyniku rozdzielenia ładunków po obu stronach membrany:
A 3 = F.E.
gdzie F- liczba ładunków elektrycznych w molu substancji (stała Faradaya); mi- wyrażona w woltach różnica potencjałów elektrycznych wynikająca z rozdziału ładunków po obu stronach błony komórkowej.
Łącząc wzory, łatwo jest uzyskać dobrze znane równanie Nernsta, czyli potencjał równowagi (dyfuzji):

Podstawiając wartości rzeczywiste w temperaturze pokojowej do wzoru równowagi sił, otrzymujemy rzeczywisty ładunek membrany, który nazywamy potencjałem membrany. Zwykle waha się w różnych komórkach od -60 do -90 mV. Bezpośrednie pomiary ładunków błony komórek nerwowych dały identyczne wyniki. Typowa komórka nerwowa kręgowca ma stały potencjał błonowy około -75 mV. Należy zauważyć, że jony potasu, sodu i chloru są ruchliwe, dlatego przy dokładnym obliczaniu potencjałów błonowych należy wziąć pod uwagę inne jony.
Ładunek błony i jej zdolność do zmiany przepuszczalności jonów pod wpływem różnych przyczyn jest wyjątkowym osiągnięciem ewolucyjnym. Wiele komórek ma tę właściwość. Jednak komórki nerwowe wykorzystują go do odbierania, przekazywania i przechowywania informacji. Jeśli podczas lokalnego działania na komórkę nerwową pojawia się obszar lokalny o zmienionym ładunku, sygnał rozchodzi się wzdłuż błony komórki nerwowej. Nazywa się to zwykle pasywnym, a sam potencjał nazywany jest stopniowym. Oznacza to, że powstałe wzbudzenie lokalne rozchodzi się wzdłuż membrany elektrotonicznie, co prowadzi do jej stopniowego tłumienia. Zazwyczaj takie sygnały rozchodzą się na krótkie odległości, chociaż u stawonogów znane są neurony, które przekazują takie sygnały na przestrzeni kilkudziesięciu milimetrów. Stopniowe potencjały tworzą światłoczułe komórki siatkówki owadów i kręgowców, wiele receptorów obwodowych, a nawet neurony ruchowe mięśni ściany ciała glisty.
Wszystkie komórki na tej planecie mają potencjał spoczynkowy. Jednak w układzie nerwowym jest tylko punktem wyjścia do odbierania, przechowywania, przetwarzania i przesyłania sygnałów. Informacja jest przekazywana w komórkach nerwowych za pomocą dynamicznej zmiany ładunku błony komórkowej. Ta zmiana może być szybka lub wolna. Jeśli następuje to szybko, a zmiana ładunku błony komórkowej przekracza pewien próg, wówczas pojawia się potencjał czynnościowy. Neurony, które tworzą potencjał czynnościowy, nazywane są neuronami kolczastymi. Potencjał czynnościowy różni się od potencjału stopniowego tym, że nie zanika, gdy oddala się od swojego źródła. Powodem tego jest zdolność membrany do aktywnej propagacji lokalnego ponownego ładowania membrany, jeśli zmiana jej potencjału osiągnie określony poziom. W normalnej sytuacji błona komórki nerwowej o potencjale spoczynkowym -75 mV powinna zmienić swój ładunek na +40 mV. Taka zmiana prowadzi do powstania nietłumionego potencjału czynnościowego, który rozprzestrzenia się wzdłuż błony komórkowej. Wartość ładunku błony, od której rozpoczyna się propagacja sygnału, nazywana jest progiem. Ponieważ procesy komórek nerwowych są wystarczająco dobrze izolowane przez komórki glejowe, a potencjał rozprzestrzenia się wzdłuż błony, jego propagacja jest spazmatyczna (słoniowa). W miejscach, gdzie komórki glejowe ściśle przylegają do błon neuronalnych, przewodzenie zachodzi nagle: od jednego przejęcia Ranviera do drugiego. W węzłach membrana procesowa jest otwarta, co umożliwia propagację sygnału poprzez otwieranie i zamykanie kanałów jonów potasu i sodu.
Zasada przekazywania sygnału przez błonę komórki nerwowej jest dość prosta. Opiera się na dużej gęstości kanałów jonowych w błonie komórek nerwowych. Liczba kanałów sodowych w węzłach Ranvier może sięgać 12.000 na 1 µm 2 . Duża liczba kanałów jonowych w błonach komórek nerwowych jest podstawą propagacji potencjałów czynnościowych. Kanały sodowe i potasowe są zlokalizowane dość gęsto, co pozwala na powstawanie lokalnych przeciwprądów jonów podczas ruchu sygnału. Potencjał czynnościowy powstaje w neuronie na wzgórku aksonu i rozpoczyna się wraz z otwarciem kanałów jonowych sodu i wnikaniem sodu do komórki. Ponieważ w spoczynku błona neuronu ma wewnętrzny ładunek powierzchniowy około -70 mV, zaczyna się niewielkie przesunięcie potencjału. Ładunek membrany lokalnie maleje i stopniowo osiąga zero, a następnie do +40 mV.
Zmiana znaku ładunku błony prowadzi do zamknięcia kanałów sodowych i otwarcia kanałów potasowych. Przepływ jonów potasu przywraca pierwotny ładunek ujemny. Proces ten nazywa się krótkoterminowym odwróceniem potencjału lub potencjałem czynnościowym. Powstając w pagórku aksonu, rozprzestrzenia się wzdłuż błony komórkowej. Fala zmiany ładunku membrany porusza się z dużą prędkością, a cały proces rewersji i regeneracji trwa milisekundy. Jednak szybkości ruchu potencjałów czynnościowych różnią się znacznie. Wpływa na to stopień mielinizacji, średnica włókien nerwowych i wiele innych czynników.
Szybkości przewodzenia potencjałów czynnościowych w układzie nerwowym u różnych gatunków zwierząt różnią się dziesięciokrotnie. Największe prędkości występują w aksonach człowieka i krewetek (120–200 m/s), a najmniejsze w ukwiałach i meduzach (0,1–0,5 m/s). W rzeczywistości procesy sygnalizacyjne zachodzą w układzie nerwowym w tempie, które odzwierciedla dynamikę i długość życia organizmu. Szybkość procesów nerwowych determinuje aktywność zwierzęcia i wewnętrzną percepcję czasu.
Należy zauważyć, że przesyłane informacje są kodowane przez częstotliwość, sekwencję i czas trwania aktywności impulsu, co zapewnia wysoką dokładność przesyłanych informacji. Te sygnały informacyjne różnią się nie tylko zasadami kodowania, ale także źródłami – rodzajami komórek nerwowych. Istnieje klasyfikacja morfologiczna i fizjologiczna neuronów. Zgodnie z wiodącymi funkcjami, neurony są fizjologicznie podzielone na czuciowe (doprowadzające), motoryczne lub motoryczne (eferentne), interkalarne (asocjacyjne, łączące aferentne i odprowadzające) i neurosekrecyjne (komórki hormonalne układu nerwowego). Te typy nie są absolutnie sztywne, a każda komórka częściowo posiada wszystkie wymienione funkcje.
Z książki Mikrobiologia: notatki do wykładów autor Tkaczenko Ksenia Wiktorowna2. Budowa ściany komórkowej i błony cytoplazmatycznej Ściana komórkowa jest elastyczną, sztywną formacją o grubości 150–200 angstremów. Pełni następujące funkcje: 1) ochronne, fagocytoza; 2) regulacja ciśnienia osmotycznego; 3) receptor; 4) akceptuje
Z książki Biologia [Kompletny przewodnik po przygotowaniu się do egzaminu] autor Lerner Georgy Isaakovich Z książki Podstawy psychofizjologii autor Aleksandrow Juriń Z książki Problemy głodu terapeutycznego. Badania kliniczne i eksperymentalne [wszystkie cztery części!] autor Anokhin Petr Kuźmich1. REJESTRACJA AKTYWNOŚCI PULSU KOMÓREK NERWOWYCH Badanie aktywności komórek nerwowych, czyli neuronów, jako integralnych jednostek morfologicznych i funkcjonalnych układu nerwowego, pozostaje oczywiście podstawowym kierunkiem w psychofizjologii. Jeden ze wskaźników aktywności
Z książki Pochodzenie mózgu autor Saveliev Siergiej WiaczesławowiczCechy mobilności głównych procesów nerwowych u pacjentów z różnymi chorobami psychicznymi podczas leczenia ich głodu Yu S. NIKOLAEV, VA BRYUZGIN, VB, GURVICH
Z książki W poszukiwaniu pamięci [Powstanie nowej nauki ludzkiej psychiki] autor Kandel Eric Richard§ 9. Kontakty synaptyczne komórek nerwowych Każdy neuron jest w stanie odbierać i przekazywać informacje. Robi to w specjalnych obszarach membrany (patrz rys. I-12). Pojedyncze aksony zwykle odchodzą od ciała neuronu, wzdłuż którego neuron przekazuje potencjały czynnościowe lub
Z książki Geny i rozwój ciała autor Neifach Aleksander Aleksandrowicz§ 10. Rodzaje połączeń komórek nerwowych Komórki nerwowe łączą się w układy nerwowe na różne sposoby. W najprostszym przypadku elementy te są rozmieszczone dość równomiernie na całym ciele zwierzęcia lub na jego większości (Anderson, 1990). Równomierna dystrybucja komórek nerwowych
Z książki Mózg w polach elektromagnetycznych autor Chołodow Jurij AndriejewiczRozdział II. Pojawienie się komórek nerwowych i mózgu Przyczyną powstania układu nerwowego był niski wskaźnik pozyskiwania informacji o zewnętrznym i wewnętrznym świecie ciała z organizacją przednerwową. Jego tkanki składały się z komórek o podobnych właściwościach chemicznych, elektromagnetycznych i
Z książki Zachowanie: podejście ewolucyjne autor Kurczanow Nikołaj Anatolijewicz Z książki Chemia biologiczna autor Lelevich Władimir Walerjanowicz Z książki autora2. Płeć komórek płciowych Po dostaniu się gonocytów do gruczołów płciowych ich los zależy od tego, jakie komórki płciowe powinny utworzyć - męskie, czyli plemniki, czy żeńskie, czyli jaja. W pierwszym przypadku ich ścieżka rozwojowa nazywana jest spermatogenezą, a w drugim -
Z książki autora1. Kształt komórek Kształt komórek zależy od ich budowy wewnętrznej i właściwości błony komórkowej oraz od otoczenia - sąsiadujących komórek i powierzchni styku. Tak więc, gdy pojedyncze komórki hoduje się na szklanej powierzchni, wszystkie komórki mają tendencję do rozprzestrzeniania się na podłożu.
Z książki autoraRozdział 9. Błony i biochemia Mikroskop elektronowy wykazał, że reakcje biochemiczne w żywej komórce przebiegają przy aktywnym udziale procesów błonowych. Wniosek ten dotyczy również komórek nerwowych i glejowych oraz organelli wewnątrzkomórkowych
Z książki autora7.8. Sygnały komórek nerwowych Układ nerwowy powstaje w toku ewolucji, aby organizmy analizowały napływające informacje. W tym celu komórki nerwowe wykorzystują sygnały elektryczne i chemiczne. Istnieją dwa rodzaje sygnałów elektrycznych.Potencjał stopniowy występuje w
Z książki autoraRozdział 9
Z książki autoraRola neuroprzekaźników w przekazywaniu impulsów nerwowych Większość synaps w układzie nerwowym ssaków ma charakter chemiczny. Proces przekazywania sygnału w synapsie chemicznej odbywa się poprzez uwalnianie neuroprzekaźników z presynaptycznych zakończeń nerwowych. Do
Dział nauk przyrodniczych związany z anatomią, fizjologią, biochemią, biologią molekularną tkanki nerwowej oraz związany z zachowaniem i uczeniem się nazywa się neuronauką. Główne elementy neuronauki to neurobiologia, psychofizjologia, neurofizjologia.
Neurobiologia to ogólna nazwa nauki, która bada na wielu poziomach (od molekularnego do behawioralnego) układ nerwowy i mózg jako jego główny narząd. Specjaliści z tej dziedziny starają się wniknąć w molekularne, komórkowe, biochemiczne mechanizmy procesów nerwowych. Psychofizjologia, czyli psychobiologia, to dziedzina wiedzy o biologicznych mechanizmach zjawisk psychicznych. Jego celem jest zrozumienie, w jaki sposób z pracy mózgu, którą można przedstawić jako wyniki obiektywnych pomiarów, powstaje świat zjawisk psychicznych. Neurofizjologia bada subtelne mechanizmy pracy komórek nerwowych w mózgu, zwanych neuronami, naturę ich wzbudzania i hamowania.
W tym artykule podajemy dość szczegółowy opis mechanizmów interakcji neuronów przez synapsy i szczegółowo rozważamy substancje w nich zaangażowane. Mówi również o substancjach, które mogą wpływać na takie interakcje. Są to, jak można się domyślać, substancje psychoaktywne, a w szczególności narkotyki.
Najbardziej ogólna esencja obrazu przedstawionego w artykule jest następująca. Jednostką strukturalną i funkcjonalną mózgu jest komórka nerwowa – neuron. Ciała komórek nerwowych tworzą istotę szarą mózgu, a ich procesy, z których powstają ścieżki i nerwy, tworzą istotę białą. Tkanka nerwowa – zbiór komórek nerwowych – jest charakterystycznym, choć nie jedynym składnikiem układu nerwowego. Neurony to wyspecjalizowane komórki, które są w stanie odbierać sygnały z analizatorów (narządów zmysłów) i innych neuronów, przetwarzać je na impulsy nerwowe i przewodzić te impulsy do zakończeń nerwowych, które są w kontakcie z innymi neuronami lub komórkami narządów, które otrzymują określone polecenia z układu nerwowego . Oddziaływanie bodźca na receptory jest przekształcane przez neurony na procesy elektryczne. Jest to ogólna zasada dla każdego narządu zmysłów i wszelkich sygnałów dochodzących z zewnątrz do układu nerwowego żywej istoty: wszystkie wpływy ze świata zewnętrznego i wewnętrznego są „napisane” w języku procesów elektrycznych. Przekazywanie informacji między neuronami lub między neuronem a inną komórką (komórką receptorową narządu czuciowego, komórką mięśniową lub gruczołową itp.) odbywa się za pomocą synaps - specjalnych cienkich szczelin między stykającymi się komórkami. Synapsy kończą procesy neuronów - aksonów i dendrytów (mogą również znajdować się bezpośrednio na ciele neuronu). Impuls jest przekazywany wzdłuż aksonu lub dendrytu neuronu w postaci elektrycznej (potencjał błonowy). Gdy tylko impuls dociera do końca procesu neuronowego (terminalu), potencjał błonowy powoduje ciąg zdarzeń prowadzących do uwolnienia do synapsy określonych substancji – neuroprzekaźników (lub neuroprzekaźników). Przechodzą przez szczelinę między kontaktującymi się komórkami i są absorbowane przez odpowiednie miejsca (receptory) na powierzchni neuronu (innej komórki) znajdującej się po drugiej stronie synapsy. W efekcie impuls jest przekazywany do tego neuronu lub przynajmniej jest na nim ułatwione wystąpienie impulsu (są jednak takie neuroprzekaźniki, które nie ułatwiają, ale hamują przewodzenie impulsu na komórce, która je otrzymała ).
Tak więc wprowadzanie sygnałów do neuronu odbywa się przez dendryty - główne miejsce powstawania synaps pobudzających i hamujących, które wpływają na neuron. Błona dendrytyczna reaguje na mediator, informacja jest przekazywana do ciała neuronu, gdzie jest łączona i integrowana, a na wyjściu przechodzi przez akson do jego zakończeń - końców, których błona uwalnia mediatora , jest przekazywana do następnej grupy neuronów. Propagacja impulsów przez neurony (pobudzenie) i tłumienie przekazywania impulsów (hamowanie) - te dwa przeciwnie skierowane procesy leżą u podstaw zrównoważonego, harmonijnego funkcjonowania układu nerwowego - przedmiotu neurofizjologii.
Dendryty tworzą układ rozgałęzień, który odbiera sygnały (pobudzające i hamujące), które są gromadzone w obszarze wyzwalającym (wzgórze aksonów) błony komórek nerwowych. W stanie normalnym (nieaktywnym) neuron ma ładunek ujemny. Różnica potencjałów ze środowiskiem otaczającym neuron wynosi –70 mV i jest utrzymywana przez błonę komórkową, co zapewnia obniżoną zawartość dodatnich jonów sodu w komórce i zwiększoną zawartość ujemnych jonów potasu. Jeżeli w krótkim odstępie czasu całkowity sygnał w obszarze wyzwalającym przekroczy –50 mV, to w neuronie rozwija się proces lawinowy, w którym na skutek zmian intensywności jonów sodu i potasu przepływa między ciałem neuronu a w środowisku, potencjał komórki osiąga +50 mV. Powstaje impuls („kolec”), który jest przekazywany wzdłuż pojedynczego aksonu do innych neuronów. Po wytworzeniu kolca następuje okres zwany okresem refrakcji, podczas którego komórka odzyskuje swój początkowy potencjał. Okres ten trwa do kilku ms.
Do przesyłania impulsów wzdłuż aksonu wykorzystuje się osłonkę mielinową, która co milimetr jest przerywana przez węzły zwane węzłami Ranviera. Impulsy są przesyłane w wstrząsach od węzła do węzła, co zapewnia, oprócz minimalizacji kosztów energii, znaczną prędkość transmisji sygnału, która może osiągnąć 100 m/s.
Akson na końcu ma wiele gałęzi, z których każda kończy się synapsą. Wszystkie synapsy w ośrodkowym i obwodowym układzie nerwowym składają się z trzech elementów: błony presynaptycznej, błony postsynaptycznej i szczeliny synaptycznej. Synaptyczne zakończenie aksonu presynaptycznego jest oddzielone od dendrytu postsynaptycznego wąską szczeliną synaptyczną.
Mediatory są syntetyzowane w błonie presynaptycznej. Depolaryzacja błony presynaptycznej: nadchodzący potencjał czynnościowy prowadzi do uwolnienia mediatora do szczeliny synaptycznej i jego wpływu na błonę postsynaptyczną. W takim przypadku mediator może mieć zarówno działanie pobudzające, jak i hamujące. Zakończenie nerwowe i błona postsynaptyczna w synapsie nerwowo-mięśniowej nazywane są płytką końcową lub motoryczną.
Transmisja sygnału w synapsie zachodzi chemicznie. Synapsy zawierają dużą liczbę bąbelków specjalnej substancji chemicznej (pęcherzyków), które pod wpływem napływającego sygnału pękają i wypełniają swoją zawartością szczelinę synaptyczną - neuroprzekaźniki. W szczelinie synaptycznej substancja pełni rolę przekaźnika, który przekazuje wzbudzenie z aksonu neuronu presynaptycznego do dendrytu neuronu postsynaptycznego. W ten sposób neuron postsynaptyczny odbiera sygnały przez rozgałęziony układ swoich dendrytów i przy wystarczającej łącznej mocy tych sygnałów reaguje wzbudzaniem i przekazywaniem sygnałów do innych neuronów. Siła połączenia synaptycznego nie jest stała. W procesie życia połączenie synaptyczne może się wzmacniać lub osłabiać, co odzwierciedla procesy uczenia się i adaptacji.
W zależności od roli, jaką odgrywają neurony w sieci neuronowej, istnieją neurony efektorowe, które przekazują sygnał bezpośrednio do narządów wykonawczych (na przykład mięśni), interneurony (lub interneurony), które służą jako ogniwa pośrednie wzdłuż ścieżki transmisji sygnału, oraz neurony receptorowe, które odbierają sygnały z narządów zmysłów.
Należy zauważyć, że neuron nie jest elementem binarnym, który jest bierny lub wzbudzony. Poziom aktywności neuronów jest kodowany przez częstotliwość sekwencji impulsów (modulacja częstotliwości impulsów), która może wahać się od 1 do 100 Hz.
Neuron
Neuron (komórka nerwowa) to strukturalna i funkcjonalna jednostka układu nerwowego. Komórka neuronowa ma złożoną strukturę, jest wysoce wyspecjalizowana i składa się z ciała (soma) o średnicy od 3 do 100 mikronów, zawierającego jądro i inne organelle oraz procesy. (Strukturę komórki można znaleźć w rozdziale „Dodatek: struktura komórki eukariotycznej.”)
Organelle neuronowe
Soma neuronów zawiera jądro neuronu i jąderko neuronu, a także dobrze rozwinięty aparat biosyntezy, który wytwarza składniki błon, syntetyzuje enzymy i inne związki chemiczne niezbędne do wyspecjalizowanych funkcji komórek nerwowych. Aparat biosyntezy w neuronach obejmuje ciała Nissla - spłaszczone cysterny ziarnistej retikulum endoplazmatycznego, które ściśle przylegają do siebie, a także dobrze zdefiniowany kompleks Golgiego. Ponadto soma zawiera liczne elementy mitochondrialne i cytoszkieletu, w tym neurofilamenty i mikrotubule. W wyniku niecałkowitej degradacji składników błony powstaje pigmentowa lipofuscyna, która z wiekiem gromadzi się w wielu neuronach. Niektóre grupy neuronów w pniu mózgu (np. neurony istoty czarnej i sina) zawierają barwnik melatoninę.



Ryż. Schematyczne przedstawienie komórki nerwu ruchowego: 1 - jądro; 2 - jąderko; 3 - koperta jądrowa; 4 - ziarnista retikulum endoplazmatyczne; 5 - kompleks Golgiego; 6 - dendryty; 7 - akson; 8 - pagórek aksonów; 9 - początkowy segment aksonu; 10 - osłonka mielinowa; 11 - telodendrium; 12 - przedterminowe i (13) końcowe części włókna nerwowego (akson); 14 - część presynaptyczna.
Jąderko zawiera dużą ilość RNA pokrytego cienką warstwą DNA. Istnieje pewien związek między rozwojem jąderka i substancji bazofilowej w ontogenezie a powstawaniem pierwotnych reakcji behawioralnych u ludzi. Wynika to z faktu, że aktywność neuronów, nawiązywanie kontaktów z innymi neuronami zależy od akumulacji w nich substancji bazofilowych.
Jądro neuronu otoczone jest porowatą dwuwarstwową błoną. Przez pory następuje wymiana między nukleoplazmą a cytoplazmą. Kiedy neuron jest aktywowany, jądro zwiększa swoją powierzchnię dzięki występom, co wzmacnia relacje jądrowo-plazmatyczne, które stymulują funkcje komórki nerwowej. Jądro neuronu zawiera materiał genetyczny. Aparat genetyczny zapewnia różnicowanie, ostateczną formę komórki, a także typowe dla tej komórki połączenia. Inną istotną funkcją jądra jest regulacja syntezy białek neuronowych przez całe jego życie.
Neurony mają wiele cech wspólnych dla wszystkich komórek ciała. Niezależnie od lokalizacji i funkcji, każdy neuron, jak każda inna komórka, ma: błona plazmatyczna określenie granic pojedynczej komórki. Oddziela cytoplazmę komórki z licznymi zawartymi w niej organellami (jądro, aparat Golgiego, mitochondria itp.) Od płynu pozakomórkowego. Kiedy neuron komunikuje się z innymi neuronami lub wyczuwa zmiany w lokalnym środowisku, robi to poprzez błonę plazmatyczną i jej maszynerię molekularną.
Błona pełni funkcję barierową, chroniącą komórkę przed wnikaniem obcych substancji, oraz transportową, zapewniającą dostanie się do komórki substancji niezbędnych do jej życiowej aktywności. Poprzez błonę komórkową reguluje się nie tylko wchodzenie i wychodzenie substancji, ale także wymiana informacji między komórką a środowiskiem zewnątrzkomórkowym. Błony komórek nerwowych zawierają wiele receptorów, których aktywacja prowadzi do wzrostu wewnątrzkomórkowego stężenia cyklicznego adenozynomonofosforanu (cAMP) i cyklicznego guanozynomonofosforanu (cGMP), które regulują metabolizm komórkowy.
Ważne jest, aby jasno zrozumieć, że wielowarstwowa błona, która otacza ciało (soma) neuronu, zapewnia tworzenie i propagację potencjału elektrotonicznego do wzgórka aksonu. Neurony są w stanie pełnić swoją funkcję informacyjną głównie dzięki temu, że ich błona ma szczególne właściwości. Błona neuronu ma grubość 6 nm i składa się z dwóch warstw cząsteczek lipidów, które swoimi hydrofilowymi końcami zwrócone są w kierunku fazy wodnej: jedna warstwa cząsteczek jest skierowana do wewnątrz, druga na zewnątrz komórki. Końce hydrofobowe są zwrócone do siebie - wewnątrz membrany. Białka błonowe są wbudowane w podwójną warstwę lipidową i pełnią kilka funkcji: białka „pompujące” zapewniają ruch jonów i cząsteczek wbrew gradientowi stężenia w komórce; białka osadzone w kanałach zapewniają selektywną przepuszczalność błony; białka receptorowe rozpoznają pożądane cząsteczki i mocują je na błonie; enzymy znajdujące się na błonie ułatwiają przepływ reakcji chemicznych na powierzchni neuronu. W niektórych przypadkach to samo białko może być zarówno receptorem, enzymem, jak i „pompą”.
W przeciwieństwie do większości innych komórek ciała dojrzałe neurony nie mogą się dzielić, a genetycznie zdeterminowane produkty każdego neuronu muszą zapewniać zachowanie i zmianę jego funkcji przez całe życie.
Organelle
Nazywa się wszystko wewnątrz błony plazmatycznej (z wyjątkiem jądra) cytoplazma. Zawiera organelle cytoplazmatyczne niezbędne do istnienia neuronu i wykonywania jego pracy.
Mitochondria dostarczają komórce energii, wykorzystując cukier i tlen do syntezy specjalnych cząsteczek o wysokiej energii, które są w razie potrzeby zużywane przez komórkę. Odgrywają ważną rolę w oddychaniu komórkowym. Większość z nich znajduje się w najbardziej aktywnych częściach neuronu: wzgórku aksonu, w okolicy synaps. Wraz z aktywną aktywnością neuronu wzrasta liczba mitochondriów.
mikrotubule- cienkie struktury podporowe wewnątrz aksonu - pomagają neuronowi utrzymać określony kształt. Sieć wewnętrznych kanalików błonowych, przez które komórka rozprowadza produkty niezbędne do jej funkcjonowania, nazywa się retikuloma endoplazmatyczna.
Istnieją dwa rodzaje retikulum endoplazmatycznego. Błony siateczki „szorstkiej” lub ziarnistej są usiane rybosomy niezbędne dla komórki do syntezy białek, które wydziela. Obfitość elementów siateczki szorstkiej w cytoplazmie neuronów charakteryzuje je jako komórki o bardzo intensywnej aktywności wydzielniczej. Białka przeznaczone tylko do użytku wewnątrzkomórkowego są syntetyzowane na licznych rybosomach, które nie są przyłączone do błon retikulum, ale znajdują się w cytoplazmie w stanie wolnym. Inny rodzaj retikulum endoplazmatycznego nazywa się „gładkim”
Organelle zbudowane z gładkich błon retikulum pakują produkty wydzielnicze w „worki” takich błon, aby następnie przenieść je na powierzchnię komórki, gdzie są wydalane. Nazywa się również gładką siateczkę endoplazmatyczną Aparat Golgiego, nazwany na cześć Włocha Emilio Golgiego, który jako pierwszy opracował metodę barwienia tej wewnętrznej struktury, która umożliwiła jej badanie mikroskopowe. W centrum cytoplazmy znajduje się Jądro komórkowe. Tutaj neurony, podobnie jak wszystkie komórki z jądrami, zawierają informację genetyczną zakodowaną w chemicznej strukturze genów. Zgodnie z tymi informacjami w pełni uformowana komórka syntetyzuje określone substancje, które określają formę, chemię i funkcję tej komórki.
Rybosomy znajdują się z reguły w pobliżu jądra i przeprowadzają syntezę białek na matrycach tRNA. Rybosomy neuronów stykają się z retikulum endoplazmatycznym kompleksu blaszkowatego i tworzą substancję bazofilową.
Substancja bazofilowa (substancja Nissla, substancja tigroid, tigroid) jest strukturą rurkową pokrytą drobnymi ziarnami, zawiera RNA i bierze udział w syntezie składników białkowych komórki. Przedłużone pobudzenie neuronu prowadzi do zaniku substancji bazofilowej w komórce, a tym samym do zaprzestania syntezy określonego białka. U noworodków neurony płata czołowego kory mózgowej nie mają substancji bazofilowej. Jednocześnie w strukturach zapewniających istotne odruchy - rdzeniu kręgowym, pniu mózgu, neurony zawierają dużą ilość substancji bazofilowej. Porusza się prądem aksoplazmatycznym z somy komórki do aksonu.
Kompleks płytkowy (aparat Golgiego)- organelle neuronu otaczające jądro w postaci sieci. Kompleks płytkowy bierze udział w syntezie neurosekrecji i innych biologicznie czynnych związków komórki. Aparat Golgiego to system błon wewnątrzkomórkowych, z których każda jest serią spłaszczonych cystern i pęcherzyków wydzielniczych. Ten system błon cytoplazmatycznych nazywany jest siateczką agranularną ze względu na brak rybosomów przyczepionych do jego cystern i pęcherzyków.
Kompleks płytkowy bierze udział w transporcie niektórych substancji z komórki, w szczególności białek i polisacharydów. Znaczna część białek syntetyzowanych w rybosomach na błonach ziarnistej retikulum endoplazmatycznego, po wejściu do kompleksu blaszkowatego, jest przekształcana w glikoproteiny, które są pakowane w pęcherzyki wydzielnicze, a następnie uwalniane do środowiska zewnątrzkomórkowego. Wskazuje to na obecność ścisłego połączenia między kompleksem blaszkowatym a błonami ziarnistej retikulum endoplazmatycznego.
Lizosomy a ich enzymy są pęcherzykami ograniczonymi prostą błoną i zapewniającymi fagocytozę komórki. Zawierają zestaw enzymów hydrolitycznych zdolnych do hydrolizowania substancji, które dostały się do komórki. W przypadku śmierci komórki dochodzi do pęknięcia błony lizosomalnej i rozpoczyna się proces autolizy - uwalniane do cytoplazmy hydrolazy rozkładają białka, kwasy nukleinowe i polisacharydy. Prawidłowo funkcjonująca komórka jest niezawodnie chroniona przez błonę lizosomalną przed działaniem hydrolaz zawartych w lizosomach.
Pigmenty neuronów - melanina i lipofuscyna znajdują się w neuronach istoty czarnej śródmózgowia, w jądrach nerwu błędnego oraz w komórkach układu współczulnego.
Neurotubule penetrują somę neuronu i biorą udział w przechowywaniu i przekazywaniu informacji.
procesy nerwowe
Neurony różnią się znacznie kształtem, formowanymi połączeniami i sposobem funkcjonowania. Najbardziej oczywistą różnicą między neuronami a innymi komórkami jest różnorodność ich rozmiarów i kształtów. Większość komórek ciała ma kształt kulisty, sześcienny lub płytkowy. Neurony natomiast charakteryzują się nieregularnymi zarysami: mają wyrostki, często liczne i rozgałęzione. Procesy te to żywe „druty”, za pomocą których obwody neuronowe.
Istnieją dwa rodzaje procesów: dendryty i aksony. Neuron ma rozwinięty cytoszkielet, który wnika w jego procesy. Cytoszkielet zachowuje kształt komórki, jego nici służą jako „szyny” do transportu organelli i substancji upakowanych w pęcherzykach błonowych, np. neuroprzekaźników. W ciele neuronu ujawnia się rozwinięty aparat syntetyczny.
akson(Grecka „oś”) - zwykle długi proces przystosowany do prowadzenia wzbudzenia z ciała neuronu do dendrytów innego neuronu. Końce aksonu rozgałęziają się silnie, jeden akson może kontaktować się z 5000 komórek nerwowych i tworzyć do 10 000 kontaktów.
Wyspecjalizowany obszar ciała komórki (zwykle soma, ale czasami dendryt), z którego odchodzi akson, nazywa się pagórkiem aksonu. Akson i wzgórek aksonu różnią się od somy i proksymalnych części dendrytów tym, że nie mają ziarnistej retikulum endoplazmatycznego, wolnych rybosomów i kompleksu Golgiego.
Dendryty- z reguły krótkie i silnie rozgałęzione procesy, które odbierają sygnał i służą jako główne miejsce powstawania synaps pobudzających i hamujących, które wpływają na neuron (różne neurony mają inny stosunek długości aksonu i dendrytów). Termin ten, wywodzący się z greckiego słowa dendron, „drzewo”, oznacza, że mają one kształt przypominający drzewo.
Dendryty mają wyrostki boczne (kolce), które zwiększają ich powierzchnię i są miejscami największego kontaktu z innymi neuronami.
Na dendrytach i na powierzchni centralnej części neuronu, otaczającej jądro (i zwanej perikaryonem lub ciałem komórkowym), znajdują się synapsy wejściowe ( połączenia neuronowe ) utworzone przez aksony innych neuronów. Dzięki temu każdy neuron okazuje się być ogniwem jednej lub drugiej sieci neuronowej.
Neuron może mieć kilka dendrytów i zwykle tylko jeden akson. Jeden neuron może mieć połączenia z wieloma (do 20 tys.) innymi neuronami. Dendryty nie mają osłonki mielinowej, podczas gdy aksony mają. Miejscem generowania wzbudzenia w większości neuronów jest wzgórek aksonu - formacja w miejscu, w którym akson opuszcza ciało. We wszystkich neuronach strefa ta nazywana jest strefą wyzwalającą.
Impulsy nerwowe rozchodzą się wzdłuż procesów neuronów i są przekazywane przez synapsy (zwykle od zakończenia aksonu do somy lub dendrytu następnego neuronu). Pojawienie się i propagacja impulsu nerwowego oraz jego transmisja synaptyczna są ściśle związane ze zjawiskami elektrycznymi na błonie komórkowej neuronu.
Na podstawie liczby i rozmieszczenia dendrytów i aksonów neurony dzielą się na neurony jednobiegunowe, neurony pseudounipolarne, neurony dwubiegunowe i neurony wielobiegunowe (wiele pni dendrytycznych, zwykle odprowadzających).
Według pozycji w łuku odruchowym neurony aferentne (neurony wrażliwe), neurony odprowadzające (niektóre z nich nazywane są neuronami ruchowymi, czasami nie jest to bardzo dokładna nazwa dotyczy całej grupy odprowadzających) i interneurony (neurony interkalarne lub asocjacyjne; to grupa neuronów komunikuje się między neuronami odprowadzającymi i doprowadzającymi).
Neurony aferentne (czułe, czuciowe) - przenoszą informację o odczuciu (impulsie) z powierzchni ciała i narządów wewnętrznych do mózgu.
Neurony odprowadzające (neurony ruchowe) - przewodzą impuls („polecenie”) z mózgu i rdzenia kręgowego do wszystkich pracujących narządów.
Neurony interkalarne (asocjacyjne) - analizują informacje i podejmują decyzje.
Neuron interkalarny przekazuje pobudzenie z neuronu doprowadzającego (wrażliwego) do odprowadzającego, znajdującego się w OUN. Ciała neuronów odprowadzających znajdują się w ośrodkowym układzie nerwowym lub na obwodzie - w węzłach współczulnych, przywspółczulnych. Aksony tych komórek kontynuują w postaci włókien nerwowych do organów roboczych (dobrowolnie - szkieletowe i mimowolne - mięśnie gładkie, gruczoły).
Neuron wykonuje w organizmie trudną i delikatną pracę, dlatego należy go uwolnić od codziennej, prostej pracy (odżywianie, usuwanie toksyn, ochrona przed uszkodzeniami mechanicznymi), którą wykonują komórki neurogleju (mikroglej, oligodendroglej i astroglej). Komórki mikrogleju biorą udział w tworzeniu opon mózgowych, komórki oligodendrogleju biorą udział w tworzeniu błon (osłonek mielinowych) wokół poszczególnych procesów komórek nerwowych.
Komórki glejowe
Oprócz neuronów tkanka nerwowa zawiera jeszcze inny rodzaj komórek. Komórki glejowe otaczają komórki nerwowe i odgrywają rolę wspierającą. Pełnią funkcje podtrzymujące i ochronne i nazywane są komórkami glejowymi lub glejami. Ich liczba jest 10 razy większa niż neuronów i zajmują połowę objętości OUN. Glia pełni nie tylko funkcje wspomagające, ale także zapewnia różnorodne procesy metaboliczne w tkance nerwowej, uczestniczy w tworzeniu osłonki mielinowej oraz przyczynia się do odbudowy tkanki nerwowej po urazach i infekcjach.
Wszystkie komórki neurogleju dzielą się na dwa genetycznie różne typy: gliocyty (makroglia), mikroglej.
Makrogleje ośrodkowego układu nerwowego obejmują ependymocyty, astrocyty i oligodendrocyty.
Ependymocyty. Tworzą gęstą warstwę elementów komórkowych wyścielających kanał kręgowy i wszystkie komory mózgu. Pełnią funkcję proliferacyjną, wspierającą, uczestniczą w tworzeniu splotów naczyniówkowych komór mózgu. W splotach naczyniówkowych warstwa wyściółczaka oddziela płyn mózgowo-rdzeniowy od naczyń włosowatych. Komórki wyściółkowe komór mózgu działają jako bariera krew-mózg. Niektóre ependymocyty pełnią funkcję wydzielniczą, uczestnicząc w tworzeniu płynu mózgowo-rdzeniowego i uwalniając różne substancje czynne bezpośrednio do jamy komór mózgowych lub krwi. Na przykład w obszarze spoidła tylnego mózgu ependymocyty tworzą specjalny „narząd podspoidłowy”, który wydziela sekret, prawdopodobnie zaangażowany w regulację metabolizmu wody.
Astrocyty. Tworzą aparat podtrzymujący ośrodkowego układu nerwowego. Istnieją dwa rodzaje astrocytów: protoplazmatyczne i włókniste (włókniste). Pomiędzy nimi są też formy przejściowe. Astrocyty protoplazmatyczne leżą głównie w istocie szarej ośrodkowego układu nerwowego i pełnią funkcje delimitacyjne i troficzne. Astrocyty włókniste znajdują się głównie w istocie białej mózgu i razem tworzą gęstą sieć - aparat pomocniczy mózgu. Procesy astrocytowe na naczyniach krwionośnych i na powierzchni mózgu tworzą okołonaczyniowe błony graniczne gleju z ich końcowymi wydłużeniami, które odgrywają ważną rolę w metabolizmie między neuronami a układem krążenia. Komórki te przenoszą składniki odżywcze z krwi do neuronów.
Główną funkcją astrocytów jest podtrzymywanie i izolowanie neuronów od wpływów zewnętrznych, co jest niezbędne do realizacji specyficznej aktywności neuronów.
Te dwa typy astrocytów są ze sobą połączone i tworzą rozległą trójwymiarową przestrzeń, w której zanurzone są neurony. Często dzielą się, tworząc blizny w przypadku uszkodzenia ośrodkowego układu nerwowego. Oligodendrocyty zlokalizowane są w istocie szarej i białej. Są mniejsze niż astrocyty i zawierają jedno kuliste jądro. Niewielka liczba cienkich gałęzi odchodzi od ciała komórki, a sama zawiera cytoplazmę z dużą liczbą rybosomów.
Oligodendrocyty (oligodendrocyty). Jest to najliczniejsza grupa komórek neurogleju. Oligodendrocyty otaczają ciała neuronów w ośrodkowym i obwodowym układzie nerwowym, są częścią osłonek włókien nerwowych i zakończeń nerwowych. W różnych częściach układu nerwowego oligodendrocyty mają inny kształt. Badanie mikroskopem elektronowym wykazało, że pod względem gęstości cytoplazmy komórki oligodendrogleju zbliżają się do komórek nerwowych i różnią się od nich tym, że nie zawierają neurofilamentów.
Funkcjonalne znaczenie tych komórek jest bardzo zróżnicowane. Pełnią funkcję troficzną, biorąc udział w metabolizmie komórek nerwowych. Oligodendrocyty odgrywają istotną rolę w tworzeniu błon wokół procesów komórkowych, natomiast nazywane są neurolemmocytami (lemmocytami - komórkami Schwanna). W procesie degeneracji i regeneracji włókien nerwowych oligodendrocyty pełnią jeszcze jedną bardzo ważną funkcję - biorą udział w neuronofagii (od greckiego phagos - pożerającego), tj. usuwaj martwe neurony poprzez aktywne wchłanianie produktów rozpadu.
Makrogleje obwodowego układu nerwowego obejmują:
- Komórki Schwanna(lemmocyty) to wyspecjalizowane oligodendrocyty, które syntetyzują osłonkę mielinową włókien mielinowych. Różnią się one od oligodendrogleju tym, że zwykle obejmują tylko jeden odcinek pojedynczego aksonu. Długość takiego pokrycia nie przekracza 1 mm. Pomiędzy poszczególnymi komórkami Schwanna tworzą się osobliwe granice, które nazywane są węzłami Ranviera. Komórka Schwanna „owija się” wokół aksonu i tracąc cytoplazmę w części rany, tworzy z błony gęstą wielowarstwową osłonkę mielinową.
- komórki satelitarne- otoczkowanie neuronów zwojów nerwów rdzeniowych i czaszkowych, regulując mikrośrodowisko wokół tych neuronów w taki sam sposób, jak robią to astrocyty.
komórki mikrogleju zlokalizowane zarówno w istocie szarej, jak i białej, ale jest ich więcej w istocie szarej. Z każdego końca małego podłużnego ciała komórkowego zawierającego lizosomy i dobrze rozwinięty aparat Golgiego odchodzi gruby proces. Mniejsze gałęzie boczne odchodzą od wszystkich jego gałęzi. Kiedy mózg jest uszkodzony, komórki te zamieniają się w fagocyty i poruszając się za pomocą ruchu ameboidalnego, opierają się inwazji obcych cząstek.
Komórki mikrogleju są makrofagami glejowymi i pełnią funkcję ochronną, biorąc udział w różnych reakcjach w odpowiedzi na czynniki uszkadzające. W tym przypadku komórki mikrogleju najpierw zwiększają swoją objętość, a następnie dzielą się mitotycznie. Komórki mikrogleju zmienione przez podrażnienie nazywane są kulkami ziarnistymi.
Komórki glejowe otaczają komórki nerwowe iw niektórych miejscach są z nimi w bliskim kontakcie. Liczba komórek glejowych w układzie nerwowym jest w przybliżeniu o rząd wielkości większa niż liczba neuronów. Szczególną rolę w tworzeniu tzw. osłonek mielinowych aksonów odgrywają komórki glejowe. Osłonki mielinowe powstają u kręgowców w OUN w wyniku procesów oligodendrocytów, a na obwodzie – w wyniku tzw. komórek Schwanna. Komórki te otaczają aksony wielowarstwowymi „sprzęgami” mielinowymi, dzięki czemu większość aksonów jest nimi pokryta, a wąskie obszary między szponami pozostają otwarte - węzły Ranviera. Te ostatnie w takich włóknach mają szczególne znaczenie funkcjonalne.
Interakcja między glejem a neuronami. Pomiędzy neuronami a komórkami glejowymi znajdują się wzajemnie połączone szczeliny o wielkości 15-20 nm, tak zwana przestrzeń śródmiąższowa, która zajmuje 12-14% całkowitej objętości mózgu.
Podczas potencjału czynnościowego stężenie jonów potasu w przestrzeni śródmiąższowej może wzrosnąć od 3-4 mmol/l do 10 mmol/l, co może powodować znaczną depolaryzację komórek nerwowych. W wyniku aktywnego transportu jonów potasu jego pozakomórkowe stężenie może być niższe niż normalne, co powoduje hiperpolaryzację komórek nerwowych.
Komórki glejowe mają wysoką przepuszczalność dla jonów potasu. Gdy kilka komórek glejowych ulega depolaryzacji na skutek miejscowego wzrostu stężenia jonów potasu, pomiędzy zdepolaryzowanymi i niezdepolaryzowanymi komórkami pojawia się prąd, który powoduje wejście jonów potasu do zdepolaryzowanych komórek glejowych, w wyniku czego dochodzi do pozakomórkowego stężenia jonów potasu zmniejsza się. Ze względu na wysoką przepuszczalność komórek glejowych dla jonów potasu oraz połączenia elektryczne między nimi, komórki glejowe pełnią rolę bufora w przypadku wzrostu pozakomórkowego stężenia potasu.
Komórki mieliny i gleju. Wiele aksonów jest pokrytych osłonką mielinową, którą tworzy wielokrotnie zwinięta błona komórek glejowych (komórki Schwanna). Mielina składa się głównie z lipidów, co nadaje charakterystyczny wygląd istocie białej mózgu i rdzenia kręgowego. Dzięki osłonce mielinowej wzrasta szybkość przewodzenia potencjału czynnościowego wzdłuż aksonu, ponieważ jony mogą poruszać się przez błonę aksonu tylko w miejscach, które nie są pokryte mieliną - przechwyty Ranvier. Pomiędzy przecięciami impulsy są prowadzone wzdłuż osłonki mielinowej jak wzdłuż kabla elektrycznego. Ponieważ otwarcie kanału i przejście przez niego jonów zajmuje trochę czasu, wyeliminowanie ciągłego otwierania kanałów i ograniczenie ich zasięgu do niewielkich obszarów błony, które nie są pokryte mieliną, przyspiesza przewodzenie impulsów wzdłuż aksonu o około 10 razy.
Tylko część komórek glejowych bierze udział w tworzeniu osłonki mielinowej nerwów (komórki Schwanna) lub traktów nerwowych (oligodendrocyty). Znacznie liczniejsze komórki glejowe (astrocyty, mikrogliocyty) pełnią inne funkcje: tworzą szkielet tkanki nerwowej, zapewniają jej potrzeby metaboliczne oraz regenerację po urazach i infekcjach. Astrocyty znajdują się wokół neuronów, zapewniając ich mechaniczną ochronę, a ponadto dostarczają neuronom składniki odżywcze i usuwają toksyny. Komórki glejowe zapewniają również izolację niektórych neuronów od wpływu innych. Błona zwykle składa się z dwóch warstw białka, pomiędzy którymi znajduje się warstwa lipidów. W takiej membranie osadzone są różne cząstki. Niektóre z nich są cząsteczkami białka i przenikają przez błonę (białka integralne), tworząc punkty przejścia dla wielu jonów: sodu, potasu, wapnia, chloru. Są to tak zwane kanały jonowe. Inne cząsteczki są przyczepione do zewnętrznej powierzchni błony i składają się nie tylko z cząsteczek białka, ale także z polisacharydów. Są to receptory dla cząsteczek substancji biologicznie czynnych, takich jak mediatory, hormony itp.
Błona komórkowa
Błona komórkowa (plasmalemma, plazmolemma) to potrójna powłoka lipoproteinowa (tj. "białko tłuszczowe"), która oddziela komórkę od środowiska i przeprowadza kontrolowaną wymianę między komórką a jej otoczeniem.
Najważniejsze w tej definicji nie jest to, że błona oddziela komórkę od środowiska, ale po prostu łączy komórkę ze środowiskiem. Membrana jest aktywną strukturą komórki, stale pracuje.
Właściwości membrany:
- przepuszczalność
- półprzepuszczalność
- selektywna przepuszczalność
- przepuszczalność czynna
- kontrolowana przepuszczalność
- fagocytoza i pinocytoza
- egzocytoza
- obecność potencjałów elektrycznych i chemicznych
- zmiany potencjału elektrycznego
- drażliwość. Specjalne receptory molekularne znajdujące się na błonie mogą łączyć się z substancjami sygnałowymi (kontrolnymi), w wyniku czego może zmieniać się stan błony i całej komórki. Receptory molekularne wywołują reakcje biochemiczne w odpowiedzi na połączenie z nimi lagandów (substancji kontrolnych).
- katalityczna aktywność enzymatyczna. Enzymy mogą być osadzone w błonie lub związane z jej powierzchnią (zarówno wewnątrz, jak i na zewnątrz komórki) i tam realizują swoją aktywność enzymatyczną.
Jak widać, główną właściwością membrany jest jej przepuszczalność w stosunku do różnych substancji.
Funkcje błonowe. Najważniejszą rzeczą w pracy błony komórkowej jest wymiana różnych substancji między komórką a środowiskiem międzykomórkowym. Wynika to z takiej właściwości membrany jak przepuszczalność. Ponadto membrana reguluje tę wymianę, regulując jej przepuszczalność.
Inną ważną funkcją membrany jest tworzenie różnicy potencjałów elektrycznych między jej wewnętrzną i zewnętrzną stroną. Dzięki temu wewnątrz komórki występuje ujemny potencjał elektryczny - potencjał spoczynkowy.
Wymiana informacji między komórką a jej otoczeniem odbywa się również przez błonę. Specjalne receptory molekularne znajdujące się na błonie mogą wiązać się z substancjami kontrolnymi (hormonami, mediatorami, modulatorami) i wyzwalać reakcje biochemiczne w komórce, prowadzące do różnych zmian w komórce lub w jej strukturach.
Struktura membrany. Błona komórkowa ma uniwersalną trójwarstwową strukturę. Jego środkowa warstwa tłuszczu jest ciągła, a górna i dolna warstwa białkowa pokrywają ją w postaci mozaiki poszczególnych obszarów białkowych. Warstwa tłuszczowa jest podstawą, która zapewnia izolację komórki od otoczenia, izolując ją od otoczenia. Sam bardzo słabo przepuszcza substancje rozpuszczalne w wodzie, ale łatwo przepuszcza te rozpuszczalne w tłuszczach. Dlatego przepuszczalność membrany dla substancji rozpuszczalnych w wodzie (np. jonów) należy zapewnić za pomocą specjalnych struktur białkowych – transporterów i kanałów jonowych.

Ryż. Schemat trzech warstw membrany
Żywa komórka to mała „torba białkowo-tłuszczowa” wypełniona półpłynną zawartością galaretowatą, przez którą przenikają folie i rurki. Ściany tego worka tworzą podwójny film tłuszczowy (lipidowy), pokryty od wewnątrz i na zewnątrz białkami - błoną komórkową. Dlatego mówią, że błona ma strukturę trójwarstwową: białka-tłuszcze-białka. Wewnątrz komórki znajduje się również wiele podobnych błon tłuszczowych, które dzielą jej wewnętrzną przestrzeń na przedziały. Organelle komórkowe otoczone są tymi samymi błonami: jądro, mitochondria, chloroplasty. Tak więc membrana jest uniwersalną strukturą molekularną tkwiącą we wszystkich komórkach i wszystkich żywych organizmach.
Kanały jonowe błony. Tylko substancje rozpuszczalne w tłuszczach mogą dostać się do komórki przez błonowy film tłuszczowy. Są to tłuszcze, alkohole, gazy. Na przykład w erytrocytach tlen i dwutlenek węgla łatwo przenikają i wychodzą bezpośrednio przez błonę. Ale woda i substancje rozpuszczalne w wodzie (na przykład jony) po prostu nie mogą przejść przez błonę do żadnej komórki. Oznacza to, że potrzebują specjalnych otworów. Ale jeśli zrobisz dziurę w filmie tłuszczowym, natychmiast się zaciśnie. Co robić? Rozwiązanie znaleziono w przyrodzie: konieczne jest wykonanie specjalnych struktur transportujących białka i rozciągnięcie ich przez błonę. W ten sposób uzyskuje się kanały do przejścia substancji nierozpuszczalnych w tłuszczach - kanały jonowe błony komórkowej.
Tak więc, aby nadać swojej błonie dodatkowe właściwości przepuszczalności cząsteczek polarnych (jonów i wody), komórka syntetyzuje w cytoplazmie specjalne białka, które są następnie integrowane z błoną. Są one dwojakiego rodzaju: białka transportowe (na przykład transportujące ATPazy) i channeloformery (formery kanałów). Białka te są zintegrowane z podwójną warstwą tłuszczową błony i tworzą struktury transportowe w postaci transporterów lub w postaci kanałów jonowych. Różne substancje rozpuszczalne w wodzie mogą teraz przechodzić przez te struktury transportowe, które inaczej nie mogą przejść przez błonę tłuszczową.

Ryż. Błona neuronowa i kanały sodowe
Najprostszy schemat przedstawiający membranę z dwoma kanałami sodowymi, odpowiednio, otwartymi i zamkniętymi
Ogólnie rzecz biorąc, białka osadzone w błonie są również nazywane integralnymi, właśnie dlatego, że są niejako zawarte w składzie błony i przenikają ją na wskroś. Inne białka, nie integralne, tworzą jakby wyspy, które „pływają” po powierzchni błony: albo wzdłuż jej zewnętrznej powierzchni, albo wzdłuż jej wewnętrznej. W końcu każdy wie, że tłuszcz to dobry lubrykant i łatwo się po nim ślizgać.
Konstrukcje transportu membranowego- są to specjalne struktury białkowe wbudowane w błonę i zapewniające transport przezbłonowy, tj. transport substancji przez błonę.
Wiele substancji może samodzielnie przejść przez błonę komórkową dzięki prostej dyfuzji. Przemieszczają się „automatycznie” ze strefy ich zwiększonego skupienia do strefy niższego skupienia, ponieważ. mają tendencję do wyrównywania ich stężenia po obu stronach membrany. Tak zwykle zachowują się wszystkie substancje w roztworach: mają tendencję do równomiernego rozprowadzania się w całej objętości cieczy, próbując pokonać barierę dzielącą roztwór na komory. W zasadzie każda cząsteczka może przejść przez podwójną warstwę lipidową błony komórkowej, ponieważ tworzące ją cząsteczki lipidów zachowują pewną mobilność względem siebie i mogą od czasu do czasu oddalać się od siebie, przepuszczając różne substancje. Jednak tempo takiej pasywnej dyfuzji, tj. Przejście substancji przez błonę z obszaru o wyższym stężeniu do obszaru o niższym stężeniu może się znacznie różnić dla różnych substancji. Dla wielu substancji dyfuzja trwa tak długo, że można dla nich mówić o praktycznie nieprzepuszczalnej membranie. Szybkość dyfuzji różnych substancji przez błonę zależy głównie od wielkości ich cząsteczek, ich ładunku elektrycznego (polaryzacji) i ich względnej rozpuszczalności w tłuszczach.
Rozpuszczalne w tłuszczach niepolarne małe cząsteczki łatwiej i co najważniejsze pasywnie przenikają przez błonę. Tak więc małe niepolarne cząsteczki, takie jak O2, steroidy, hormony tarczycy i kwasy tłuszczowe, najłatwiej przechodzą przez błonę za pomocą prostej dyfuzji. Małe polarne, nienaładowane cząsteczki dyfundują przez warstwę lipidową nieco wolniej: CO2, NH3, H2O, etanol, mocznik. Dyfuzja glicerolu jest już znacznie wolniejsza, a glukoza praktycznie nie jest w stanie samodzielnie przejść przez błonę. Dla wszystkich naładowanych cząsteczek, niezależnie od wielkości, błona lipidowa jest praktycznie nieprzepuszczalna. Tak więc tylko substancje rozpuszczalne w tłuszczach, które mogą rozpuścić się w tłuszczowej (lipidowej) warstwie błony, mogą swobodnie przenikać przez błonę do komórki iz powrotem. Transport innych substancji przez błonę wymaga specjalnych mechanizmów. Jakie substancje należy „na siłę” przeciągać przez błonę? Są to wszystkie cząsteczki polarne, które są nierozpuszczalne w tłuszczach: cząsteczki wody, jony (elektrolity), a także większe cząsteczki składników odżywczych, takich jak glukoza i aminokwasy.
Do transportu substancji słabo zdolnych do dyfuzji przez warstwę lipidową błony do komórki wymagane są specjalne struktury transportowe.
Rodzaje membranowych struktur transportowych:
1. Kanały jonowe - specjalne pory (otwory) w błonie utworzone przez białka kanałowe, umożliwiające przechodzenie jonów przez błonę w obu kierunkach: do wewnątrz i na zewnątrz.
2. Translokazy to specjalne białka błonowe, które ułatwiają przechodzenie substancji przez błonę ze względu na ich tymczasowe wiązanie z substancją dyfundującą. Nie wymagają energii, działają dwukierunkowo w zależności od stężenia przenoszonej substancji.
3. Transportery - struktury białkowe, które siłą przeciągają określone substancje przez błonę komórkową w określonym kierunku z kosztami energii. Pompy jonowe są transporterami jonów. W zależności od sposobu, w jaki zużywają energię do swojej pracy, przenośniki można podzielić na „importowe” i „antyportowe”. Transportery objawów wykorzystują wspólny transport dwóch substancji w jednym kierunku: jedna z nich musi mieć dużą energię potencjalną, aby przejść przez błonę. Na przykład import do komórki za pomocą jonów sodu glukozy lub import jonów wapnia za pomocą jonów sodu. Transportery antyportowe (wymienniki) wykorzystują nadchodzący transport dwóch substancji o różnej potencjalnej energii dyfuzji. Tak np. działa pompa jonów sodowo-potasowych.
Tak więc przenoszenie substancji przez błonę komórkową odbywa się na różne sposoby.
Mechanizmy transportu substancji przez membranę:
1. Prosta dyfuzja substancji rozpuszczalnych w tłuszczach (hydrofobowych) przez warstwę tłuszczową membrany. Jest to proces pasywny pod działaniem gradientu (różnicy) stężenia substancji po przeciwnych stronach membrany. (Zobacz wideo: Pasywny transport membranowy).
2. Niekontrolowana dyfuzja (niekontrolowany transfer pasywny) substancji rozpuszczalnych w wodzie przez stale otwarte kanały jonowe błony.
3. Kontrolowana dyfuzja (kontrolowany transfer pasywny) substancji rozpuszczalnych w wodzie przez kontrolowane kanały jonowe membrany.
4. Aktywny transport substancji rozpuszczalnych w wodzie za pomocą specjalnych struktur transportu białek (transporterów) dzięki wykorzystaniu energii rozszczepiania ATP.
Jedną z najważniejszych struktur transportowych błony jest enzym ATPaza. ATPazy różnych typów transportują jony przez błonę. Przenoszą je zarówno w celi, jak i odwrotnie.
Nazwa ATPaza oznacza, że jest to enzym rozkładający ATP (trójfosfatazę adenozyny).
Obecnie zidentyfikowano wiele różnych typów transportowych ATPaz. Są podobne w budowie i mechanizmie działania, ale mają różne specjalizacje, tj. każdy z ich gatunków ciągnie przez błonę coś innego. Obecnie Na+/K+-ATPaza, Ca2+-ATPaza, H+-ATPaza, H+,K+-ATPaza, Mg2+-ATPaza, które zapewniają ruch jonów Na+, K+, Ca2+, H+, Mg2+ odpowiednio w izolacji lub w koniugacji, są dobrze zbadane: na przykład Na+ jest sprzężony z K+; H+ jest sprzężony z K+.
Jaka jest zasada działania ATPazy?
Enzymy te rozkładają ATP i uwalniają energię chemiczną zawartą w cząsteczkach ATP. Ta wyzwolona energia jest natychmiast zużywana na jakąś pożyteczną pracę. Transportowe ATPazy błonowe zużywają go na dostarczanie „siłą” określonej substancji na przeciwną stronę błony. Różne ATPazy wbudowane w błonę działają jako nośniki dla różnych substancji, a zatem są transporterami molekularnymi, które „przepychają” substancje przez błonę. Ten transfer nazywa się aktywnym transportem.
Na,K-ATPazę (trifosfataza sodowo-potasowo-adenozynowa) można słusznie uznać za najważniejszą ATP-azę błonową.
W błonie tworzy się Na,K-ATPaza „jonowa pompa sodowo-potasowa”, który przenosi jony Na + i K + po różnych stronach błony. Ważne jest, aby zrozumieć, że ta pompa działa jako wymiennik. Po wewnętrznej stronie błony miejsce aktywne enzymu (ATPaza) wychwytuje 3 jony sodu i wyrzuca je już na zewnętrzną stronę. A wyrzucając jony sodu, ATPaza wychwytuje 2 jony potasu z zewnątrz w ich miejsce. Enzym obraca się następnie wewnątrz komórki i przenosi jony potasu do wnętrza błony. Tam je uwalnia, a zamiast nich ponownie wychwytuje 3 jony sodu.
Jednocześnie należy pamiętać, że jako prawdziwy enzym Na,K-ATPaza dzieli ATP równolegle, pozyskując z niego energię do aktywności transportowej. Następnie cykl się powtarza.
spoczynkowy potencjał błonowy. Dlaczego musimy wiedzieć, jaki jest potencjał spoczynkowy? Co to jest „elektryczność zwierząt”? Skąd w ciele biorą się bioprądy? Jak żywa komórka w środowisku wodnym może zamienić się w „baterię elektryczną”?
Na te pytania będziemy w stanie odpowiedzieć, jeśli dowiemy się, jak komórka poprzez redystrybucję ładunków elektrycznych wytwarza dla siebie potencjał elektryczny na błonie.
Jak działa układ nerwowy? Gdzie to wszystko się zaczyna? Skąd pochodzi elektryczność impulsów nerwowych?
Możemy również odpowiedzieć na te pytania, jeśli dowiemy się, jak komórka nerwowa wytwarza dla siebie potencjał elektryczny na błonie.
Zatem zrozumienie, jak działa układ nerwowy, zaczyna się od zrozumienia, jak działa pojedyncza komórka nerwowa, neuron.
A praca neuronu z impulsami nerwowymi opiera się na redystrybucji ładunków elektrycznych na jego błonie i zmianie wielkości potencjałów elektrycznych. Ale aby potencjał się zmienił, musisz go najpierw mieć. Można więc powiedzieć, że neuron przygotowując się do pracy nerwowej, wytwarza na swojej błonie potencjał elektryczny jako okazję do takiej pracy.
Tak więc naszym pierwszym krokiem do zbadania funkcjonowania układu nerwowego jest zrozumienie, w jaki sposób ładunki elektryczne poruszają się po komórkach nerwowych i jak to tworzy potencjał elektryczny na błonie. To właśnie zrobimy i ten proces pojawienia się potencjału elektrycznego w neuronach nazwiemy - tworzeniem potencjału spoczynkowego.
Normalnie, gdy komórka jest gotowa do pracy, ma już ładunek elektryczny na powierzchni membrany. Nazywa się to spoczynkowym potencjałem błonowym.
potencjał spoczynkowy- jest to różnica potencjałów elektrycznych pomiędzy wewnętrzną i zewnętrzną stroną błony, gdy komórka znajduje się w stanie fizjologicznego spoczynku. Jego średnia wartość to -70 mV (miliwolty).
„Potencjał” jest szansą, jest pokrewny koncepcji „potencji”. Potencjał elektryczny membrany to jej zdolność do przenoszenia ładunków elektrycznych, dodatnich lub ujemnych. W roli ładunków pełnią naładowane cząsteczki chemiczne – jony sodu i potasu, a także wapnia i chloru. Spośród nich tylko jony chlorkowe są naładowane ujemnie (–), podczas gdy pozostałe są naładowane dodatnio (+).
Mając potencjał elektryczny, membrana może przenosić powyższe naładowane jony do lub z komórki.
Ważne jest, aby zrozumieć, że w układzie nerwowym ładunki elektryczne są wytwarzane nie przez elektrony, jak w przypadku drutów metalowych, ale przez jony - cząsteczki chemiczne, które mają ładunek elektryczny. Prąd elektryczny w ciele i jego komórkach to przepływ jonów, a nie elektronów, jak w przewodach. Należy również zauważyć, że ładunek membrany jest mierzony od wewnątrz ogniwa, a nie od zewnątrz.
Oznacza to, że jony naładowane dodatnio będą dominować na zewnątrz wokół ogniwa, a jony naładowane ujemnie będą przeważać wewnątrz. Można powiedzieć, że wewnątrz komórki jest elektroujemna.
Istotą potencjału spoczynkowego jest przewaga ujemnych ładunków elektrycznych w postaci anionów na wewnętrznej stronie błony oraz brak dodatnich ładunków elektrycznych w postaci kationów, które są skoncentrowane na jej zewnątrz, a nie wewnątrz.
Ten stan rzeczy jest osiągany przez trzy zjawiska: (1) zachowanie membrany, (2) zachowanie dodatnich jonów potasu i sodu oraz (3) związek między siłą chemiczną i elektryczną.
1. Zachowanie membrany. W zachowaniu membrany dla potencjału spoczynkowego ważne są trzy procesy:
1) Wymiana wewnętrznych jonów sodu na zewnętrzne jony potasu. Wymiana odbywa się za pomocą specjalnych konstrukcji transportowych membrany: pomp jonitowych. W ten sposób błona przesyca komórkę potasem, ale zuboża sód.
2) Otwarte kanały jonów potasowych. Za ich pośrednictwem potas może zarówno przedostawać się do komórki, jak i ją opuszczać. W zasadzie wychodzi.
3) Zamknięte kanały jonów sodowych. Z tego powodu sód usunięty z ogniwa pompami wymiennymi nie może do niej wrócić. Kanały sodowe otwierają się tylko w specjalnych warunkach - a wtedy potencjał spoczynkowy zostaje zaburzony i przesunięty w kierunku zera. Nazywa się depolaryzacja membrany, tj. zmniejszająca się polaryzacja.
2. Zachowanie się jonów potasu i sodu. Jony potasu i sodu przemieszczają się przez błonę na różne sposoby:
1) Za pomocą pomp jonowymiennych potas jest wciągany do komórki, a sód jest wydalany z komórki.
2) Poprzez stale otwarte kanały potasowe, potas opuszcza komórkę, ale może też przez nie wracać do niej.
3) Sód „chce” dostać się do celi, ale „nie może”, ponieważ kanały są dla niego zamknięte.
3. Stosunek sił chemicznych i elektrycznych.
W stosunku do jonów potasu ustala się równowaga między siłami chemicznymi i elektrycznymi na poziomie - 70 mV.
1) Siła chemiczna wypycha potas z komórki, ale ma tendencję do wciągania do niej sodu.
2) Siła elektryczna ma tendencję do wciągania dodatnio naładowanych jonów (zarówno sodu, jak i potasu) do komórki.
Powstawanie potencjału spoczynkowego. Nasze komórki są tylko dodatnie na zewnątrz, ale wewnątrz są bardzo ujemne, aw nich występuje nadmiar cząstek ujemnych - anionów i brak cząstek dodatnich - kationów. Wewnętrzna elektroujemność ogniwa nie powstaje z powodu pojawienia się dodatkowych cząstek ujemnych (anionów), ale przeciwnie, z powodu utraty pewnej ilości cząstek dodatnich (kationów). I dlatego istotą naszej opowieści nie będzie to, że wyjaśnimy, skąd w komórce biorą się ujemne cząstki, ale to, że wyjaśnimy, w jaki sposób w neuronach uzyskuje się deficyt dodatnio naładowanych jonów – kationów.
Skąd dodatnio naładowane cząstki wychodzą z ogniwa? Przypomnij sobie, że są to jony sodu - Na + i potasu - K +.
Chodzi o to, że w błonie komórki nerwowej stale pracują pompy wymienne utworzone przez specjalne białka wbudowane w błonę. Co oni robią? Zmieniają „własny” sód komórki na zewnętrzny „obcy” potas. Z tego powodu w komórce kończy się brak sodu, który trafił na wymianę. W tym samym czasie komórka jest przepełniona jonami potasu, które te pompy molekularne wciągnęły do niej.
Aby ułatwić zapamiętanie, możesz w przenośni powiedzieć: „Komórka kocha potas!” (Choć prawdziwa miłość nie wchodzi w rachubę!) Dlatego wciąga w siebie potas, mimo że jest już nim pełen. Dlatego nieopłacalnie zamienia go na sód, dając 3 jony sodu na 2 jony potasu. Dlatego na tę wymianę zużywa energię ATP. A jak wydać! Na pracę pomp sodowo-potasowych można wydać do 70% całego zużycia energii neuronów. To właśnie robi miłość, nawet jeśli nie jest prawdziwa!
Nawiasem mówiąc, ciekawe jest to, że komórka nie rodzi się z potencjałem spoczynkowym w swojej gotowej postaci. Na przykład podczas różnicowania i fuzji mioblastów potencjał ich błony zmienia się od –10 do –70 mV, tj. ich błona staje się bardziej elektroujemna, polaryzuje się podczas różnicowania.
Mówiąc obrazowo, możemy powiedzieć, że tworząc potencjał spoczynkowy, komórka jest „naładowana miłością” do potasu, a potas miłością do wolności. Co dziwne, ale rezultatem tych dwóch rodzajów miłości jest pustka!
To pustka tworzy w komórce ujemny ładunek elektryczny - potencjał spoczynkowy. Dokładniej, ujemny potencjał tworzą puste przestrzenie pozostawione po potasie, który uciekł z komórki.
Pompa wymiany jonów sodowo-potasowych stwarza trzy potencjały (możliwości):
1. Potencjał elektryczny – zdolność do wciągania dodatnio naładowanych cząstek (jonów) do wnętrza ogniwa.
2. Potencjał jonowy sodu - zdolność do wciągania jonów sodu do komórki (i jonów sodu, a nie żadnych innych).
3. Potencjał jonowy potasu - zdolność wypychania jonów potasu z komórki (i to potas, a nie wszystkie inne).
Tak więc wynik działania membranowych pomp jonowymiennych jest następujący:
1. Niedobór sodu (Na+) w komórce.
2. Nadmiar potasu (K+) w komórce.
Można powiedzieć tak: pompy jonowe błony wytwarzają różnicę w stężeniach jonów lub gradient stężenia (różnicę) między środowiskiem wewnątrzkomórkowym i zewnątrzkomórkowym.
To właśnie z powodu wynikającego z tego niedoboru sodu ten sód będzie teraz „wpełzał” do komórki z zewnątrz. Tak zawsze zachowują się substancje: mają tendencję do wyrównywania swojego stężenia w całej objętości roztworu. Jednocześnie w komórce uzyskano nadmiar jonów potasu w porównaniu ze środowiskiem zewnętrznym. Ponieważ pompy membranowe wpompowywały go do komórki. I stara się wyrównać swoją koncentrację wewnątrz i na zewnątrz, dlatego stara się wydostać z klatki.
Tutaj również ważne jest, aby zrozumieć, że jony sodu i potasu niejako „nie zauważają” siebie nawzajem, reagują tylko „na siebie”. Tych. sód reaguje na stężenie sodu, ale „nie zwraca uwagi” na to, ile potasu jest w pobliżu. Odwrotnie, potas reaguje tylko na stężenie potasu i „nie zauważa” sodu. Okazuje się, że aby zrozumieć zachowanie jonów w komórce, konieczne jest oddzielne porównanie stężeń jonów sodu i potasu. Tych. konieczne jest oddzielne porównywanie stężenia sodu wewnątrz i na zewnątrz komórki oraz osobno stężenia potasu wewnątrz i na zewnątrz komórki, ale nie ma sensu porównywać sodu z potasem, jak to często robi się w podręcznikach.
Zgodnie z prawem wyrównywania stężeń, które działa w roztworach, sód „chce” dostać się do komórki z zewnątrz. Ale nie może, ponieważ membrana w swoim normalnym stanie nie przechodzi przez nią dobrze. Wchodzi trochę i komórka ponownie natychmiast wymienia go na zewnętrzny potas. Dlatego sodu w neuronach zawsze brakuje. Ale potas po prostu może łatwo wydostać się z komórki! Klatka jest nim pełna, a ona nie może go zatrzymać. Tak więc wychodzi przez specjalne dziury białkowe w błonie (kanały jonowe).
A teraz - najważniejsza rzecz. Musimy przejść od ruchu cząstek chemicznych do ruchu ładunków elektrycznych.
Potas jest naładowany ładunkiem dodatnim, dlatego opuszczając ogniwo pobiera z niej nie tylko siebie, ale także ładunki dodatnie. W ich miejsce w ogniwie pozostają ładunki ujemne. To jest potencjał spoczynkowy błony.
Spoczynkowy potencjał błonowy to deficyt dodatnich ładunków wewnątrz komórki, powstały w wyniku wycieku dodatnich jonów potasu z komórki.
Potencjał spoczynkowy jest ujemny od strony komórki i składa się niejako z dwóch części.
1. Pierwsza część to około -10mV, które uzyskuje się z nierównobocznej pracy pompy wymiennika membranowego (w końcu więcej wypompowuje Na+ niż pompuje z powrotem z K+).
2. Druga część to potas wyciekający z ogniwa przez cały czas, wyciągając ładunki dodatnie z ogniwa. Zapewnia większość potencjału błonowego, obniżając go do -70 miliwoltów.
Potas przestanie opuszczać ogniwo (a dokładniej jego wejście i wyjście będą sobie równe) tylko wtedy, gdy poziom elektroujemności ogniwa wyniesie –90mV. Jest to jednak utrudnione przez sód stale przeciekający do komórki, który ciągnie za sobą jego dodatnie ładunki. A komórka utrzymuje stan równowagi na poziomie –70mV.
Chodzi więc o membranową pompę sodowo-potasową, a następnie wypływ „nadmiaru” potasu z ogniwa. Z powodu utraty ładunków dodatnich podczas tego wycieku wzrasta elektroujemność wewnątrz ogniwa. To właśnie ten „potencjał spoczynkowy błony”. Jest mierzony wewnątrz ogniwa i zwykle wynosi -70 mV.
Potencjał lokalny. Potencjał lokalny (LP) to lokalne, nie propagujące się wzbudzenie podprogowe, które występuje w zakresie od potencjału spoczynkowego (–70 mV) do krytycznego poziomu depolaryzacji (–50 mV).
Jeśli krytyczny poziom depolaryzacji zostanie przekroczony, lokalny potencjał przekształca się w potencjał czynnościowy i generuje impuls nerwowy.
Krytyczny poziom depolaryzacji (CDL) to poziom potencjału elektrycznego błony komórki pobudliwej, z którego potencjał lokalny przechodzi w potencjał czynnościowy. Zjawisko to polega na samonarastającym otwarciu bramkowanych napięciem kanałów jonowych dla sodu pod wpływem narastającej depolaryzacji.
FCA wynosi zwykle -50 mV, ale zmienia się w zależności od neuronu i może się zmieniać wraz ze zmianą pobudliwości neuronu. Im bliżej FRC jest do potencjału spoczynkowego (–70 mV) i odwrotnie, im bliżej potencjał spoczynkowy jest do RCC, tym bardziej pobudliwy jest neuron.
Ważne jest, aby zrozumieć, że proces lokalnego generowania potencjału rozpoczyna się wraz z otwarciem kanałów jonowych. Najważniejsze jest otwarcie kanałów jonowych. Muszą być otwarte, aby strumień jonów dostał się do komórki i wniósł do niej ładunki elektryczne. Te jonowe ładunki elektryczne powodują tylko przesunięcie potencjału elektrycznego membrany w górę lub w dół, tj. lokalny potencjał.
Jeśli kanały jonowe dla sodu (Na+) otworzą się, do komórki wnikają ładunki dodatnie wraz z jonami sodu, a ich potencjał przesuwa się w górę w kierunku zera. To jest depolaryzacja (spadek elektroujemności komórki nerwowej) i tak rodzi się ekscytujący lokalny potencjał. Można powiedzieć, że pobudzające lokalne potencjały są generowane przez kanały jonów sodowych, gdy się otwierają.
W przenośni można powiedzieć tak: „kanały otwierają się – rodzi się potencjał”.
Jeśli kanały jonowe dla chloru (Cl–) otworzą się, do ogniwa wraz z jonami chloru wpadają ładunki ujemne, a jego potencjał spada poniżej potencjału spoczynkowego. To jest hiperpolaryzacja iw ten sposób rodzi się miejscowy potencjał hamujący. Można powiedzieć, że lokalne potencjały hamujące są generowane przez kanały jonów chlorkowych.
Istnieje również inny mechanizm powstawania hamujących potencjałów lokalnych - ze względu na otwarcie dodatkowych kanałów jonowych dla potasu (K +). W tym przypadku „dodatkowe” porcje jonów potasu zaczynają przez nie opuszczać ogniwo, przenoszą ładunki dodatnie i zwiększają elektroujemność ogniwa, tj. powodować hiperpolaryzację. Można więc powiedzieć, że hamujące lokalne potencjały są generowane przez dodatkowe kanały jonowe potasu.
Jak widać, wszystko jest bardzo proste, najważniejsze jest otwarcie niezbędnych kanałów jonowych. Kanały jonowe bramkowane bodźcem otwierają się pod wpływem bodźca (bodźca). Bramkowane chemicznie kanały jonowe są otwierane przez neuroprzekaźnik (pobudzający lub hamujący). Dokładniej, w zależności od tego, na jakie kanały (sodowy, potasowy czy chlorkowy) będzie działał mediator, będzie to potencjał lokalny – pobudzający lub hamujący. A mediator, zarówno dla lokalnych potencjałów pobudzających, jak i hamujących, może być ten sam, ważne jest tutaj, które kanały jonowe zwiążą się z nim ze swoimi receptorami molekularnymi – sodowym, potasowym czy chlorkowym.
Rodzaje lokalnych potencjałów:
1. Receptor. Występuje na komórkach receptorowych lub zakończeniach receptorów neuronów pod wpływem bodźca (bodźca).
2. Generator. Występuje na czuciowych neuronach aferentnych (na ich zakończeniach nerwowych, węzłach Ranviera lub wzgórkach aksonów) pod wpływem mediatorów, które uwolniły receptory komórek czuciowych. Potencjał generatora zamienia się w potencjał czynnościowy i impuls nerwowy, gdy osiągnie krytyczny poziom depolaryzacji, tj. generuje (generuje) impuls nerwowy.
3. Pobudzający potencjał postsynaptyczny (EPSP). Występuje na błonie postsynaptycznej synapsy, tj. odzwierciedla przeniesienie pobudzenia z jednego neuronu na drugi. Powoduje depolaryzację błony. Ale zwykle potrzeba całej serii EPSP, aby narodził się impuls nerwowy, ponieważ. wielkość pojedynczego EPSP jest całkowicie niewystarczająca, aby osiągnąć krytyczny poziom depolaryzacji.
4. Hamujący potencjał postsynaptyczny (IPSP). Występuje na błonie postsynaptycznej synapsy, ale tylko jej nie pobudza, a wręcz przeciwnie, hamuje. W związku z tym ta błona postsynaptyczna jest częścią synapsy hamującej, a nie pobudzającej. IPSP powoduje hiperpolaryzację błony, tj. przesuwa potencjał spoczynkowy w dół od zera. Do tworzenia IPSP wykorzystywane są dwa mechanizmy: 1) „chlor” – otwarcie kanałów jonowych na chlor (Cl–), przez które jony chloru dostają się do komórki i zwiększają jej ujemność, 2) „potas” – otwieranie kanałów jonowych na potas ( K+), przez który wychodzą jony potasu, co również zwiększa ujemność w komórce.
5. Potencjały stymulatora są endogennymi, zbliżonymi do sinusoidalnych okresowych oscylacji potencjału błonowego o częstotliwości 0,1-10 Hz i amplitudzie 5-10 mV. Są one generowane przez neurony rozrusznika (rozruszniki) niezależnie, bez wpływu zewnętrznego. Lokalne potencjały stymulatora zapewniają, że neuron stymulatora okresowo osiąga krytyczny poziom depolaryzacji i spontanicznie (spontanicznie) generuje potencjały czynnościowe i odpowiednio impulsy nerwowe.
Potencjały lokalne powstają na receptorach czuciowych, na zakończeniach receptorów neuronów i na postsynaptycznych błonach synaps.
potencjał czynnościowy.Ściśle mówiąc, pobudzenie poruszające się wzdłuż nerwów reprezentuje impulsy nerwowe, a nie potencjały czynnościowe.
Impuls nerwowy to ruchoma fala zmian stanu błony, w tym zmian strukturalnych (otwieranie i zamykanie kanałów jonowych), chemicznych (zmiana przepływu jonów) i elektrycznych (depolaryzacja, polaryzacja dodatnia i repolaryzacja).
Ale w literaturze fizjologicznej termin „potencjał czynnościowy” jest również używany jako synonim impulsu nerwowego. Chociaż potencjał czynnościowy jest tylko elektrycznym składnikiem impulsu nerwowego.
Potencjał czynnościowy to składnik elektryczny impulsu nerwowego, który charakteryzuje zmiany ładunku elektrycznego (potencjału) w lokalnym obszarze błony podczas przechodzenia przez nią impulsu nerwowego (od -70 do +30 mV i odwrotnie ).
Potencjał czynnościowy to impuls elektryczny wywołany zmianą przepuszczalności jonów błony i związany z propagacją fali wzbudzenia przez nerwy i mięśnie. W swej istocie reprezentuje wyładowanie elektryczne - szybką krótkotrwałą zmianę potencjału na małym odcinku błony komórki pobudliwej (neuron, włókno mięśniowe lub komórkę gruczołową), w wyniku której zewnętrzna powierzchnia tego odcinka zostaje naładowana ujemnie w stosunku do sąsiednich odcinków membrany, podczas gdy jej wewnętrzna powierzchnia staje się naładowana dodatnio w stosunku do sąsiednich obszarów membrany. Potencjał czynnościowy jest fizyczną podstawą impulsu nerwowego lub mięśniowego, który odgrywa rolę sygnałową (regulacyjną).

Ryż. Komórka nerwowa i potencjał czynnościowy
Schemat rozkładu ładunków po różnych stronach błony komórki pobudliwej w stanie spoczynku (A) oraz w przypadku potencjału czynnościowego - depolaryzacji (B).
Impulsy nerwowe rodzą się na neuronach w zaledwie kilku miejscach:
- wzgórek aksonów
- receptorowy koniec dendrytu
- pierwsze przechwycenie Ranviera na dendrycie (strefa wyzwalania dendrytu)
- błona postsynaptyczna synapsy pobudzającej.
1. Wzgórze Aksonów - główny rodzic impulsów nerwowych. Wzgórze aksonu jest początkiem aksonu, gdzie zaczyna się na ciele neuronu. To właśnie wzgórek aksonu jest głównym rodzicem impulsów nerwowych na neuronie. We wszystkich innych miejscach prawdopodobieństwo narodzin impulsu nerwowego jest znacznie mniejsze. Faktem jest, że błona wzgórka aksonu ma zwiększoną wrażliwość na wzbudzenie i obniżyła krytyczny poziom depolaryzacji (CDL) w porównaniu z resztą błony. Dlatego też, gdy na błonie neuronu zaczynają się sumować liczne pobudzające potencjały postsynaptyczne (EPSP), które powstają w różnych miejscach na błonach postsynaptycznych wszystkich jego kontaktów synaptycznych, to do FEC dochodzi przede wszystkim na wzgórku aksonu. To tam ta nadprogowa depolaryzacja wzgórka otwiera wrażliwe na napięcie kanały sodowe, do których wnika przepływ jonów sodu, generując potencjał czynnościowy i impuls nerwowy.
Tak więc wzgórek aksonu jest strefą integracyjną na błonie, integruje wszystkie lokalne potencjały (pobudzający i hamujący) powstające na neuronie - a pierwszy działa, aby osiągnąć CUD, generując impuls nerwowy.
Ważne jest również, aby wziąć pod uwagę następujący fakt. Od wzgórka aksonu impuls nerwowy rozprasza się po całej błonie neuronu: zarówno wzdłuż aksonu do zakończeń presynaptycznych, jak i wzdłuż dendrytów do postsynaptycznych „początków”. Wszystkie lokalne potencjały są usuwane z błony neuronu i ze wszystkich jego synaps, ponieważ są „przerywane” przez potencjał czynnościowy z impulsu nerwowego biegnącego przez całą błonę.
2. Zakończenie receptorowe neuronu wrażliwego (dośrodkowego). Jeśli neuron ma zakończenie receptorowe, wówczas może na niego oddziaływać odpowiedni bodziec i generować na tym końcu najpierw potencjał generatora, a następnie impuls nerwowy. Kiedy potencjał generatora osiągnie KUD, na tym końcu otwierają się zależne od napięcia kanały jonów sodowych i rodzi się potencjał czynnościowy i impuls nerwowy. Impuls nerwowy biegnie wzdłuż dendrytu do ciała neuronu, a następnie wzdłuż jego aksonu do zakończeń presynaptycznych, aby przekazać wzbudzenie do następnego neuronu. Tak działają na przykład receptory bólu (nocyceptory), które są dendrytycznymi zakończeniami neuronów bólowych. Impulsy nerwowe w neuronach bólu są wychwytywane dokładnie na zakończeniach receptorów dendrytów.
3. Pierwsze przechwycenie Ranviera na dendrycie (strefa wyzwalania dendrytu). Lokalne pobudzające potencjały postsynaptyczne (EPSP) na końcach dendrytu, które powstają w odpowiedzi na wzbudzenia dochodzące do dendrytu przez synapsy, sumują się w pierwszym węźle Ranviera tego dendrytu, jeśli oczywiście jest on zmielinizowany. Istnieje odcinek membrany o zwiększonej wrażliwości na wzbudzenie (dolny próg), dlatego właśnie w tym obszarze najłatwiej jest pokonać krytyczny poziom depolaryzacji (CDL), po którym otwierają się sterowane napięciem kanały jonowe dla sodu - i powstaje potencjał czynnościowy (impuls nerwowy).
4. Błona postsynaptyczna synapsy pobudzającej. W rzadkich przypadkach EPSP w synapsie wzbudzającej może być tak silny, że dociera do CUD właśnie tam i generuje impuls nerwowy. Ale częściej jest to możliwe tylko w wyniku zsumowania kilku EPSP: albo z kilku sąsiednich synaps, które wystrzeliły jednocześnie (sumowanie przestrzenne), albo dzięki temu, że na daną synapsę dotarło kilka impulsów z rzędu (sumowanie czasowe). .
Wszystkie procesy zmian błonowych zachodzące w trakcie propagującego się wzbudzenia można rozpatrywać jako pojedynczy elektrochemiczny proces propagacji dynamicznego wzbudzenia z trzech stron, na trzech poziomach:
- Zjawiska elektryczne – rozwój potencjału czynnościowego.
- Zjawiska chemiczne - ruch przepływów jonowych.
- Zjawiska strukturalne – zachowanie kanałów jonowych.
Trzy strony procesu rozprzestrzeniania się wzbudzenia:
1. Potencjał czynnościowy (AP). Potencjał czynnościowy to nagła zmiana stałego potencjału błonowego z polaryzacji ujemnej na dodatnią i odwrotnie.
Zwykle potencjał błonowy w neuronach OUN zmienia się od –70 mV do +30 mV, a następnie ponownie powraca do swojego pierwotnego stanu, tj. do –70 mV. Jak widać, pojęcie potencjału czynnościowego charakteryzuje się zjawiskami elektrycznymi na błonie.
Na poziomie elektrycznym zmiany zaczynają się od zmiany stanu spolaryzowanego błony na depolaryzację. Po pierwsze, depolaryzacja występuje w postaci lokalnego potencjału pobudzającego. Do krytycznego poziomu depolaryzacji (około -50 mV) jest to stosunkowo prosty liniowy spadek elektroujemności proporcjonalny do siły bodźca. Ale potem zaczyna się bardziej stroma samo-wzmacniająca się depolaryzacja, która rozwija się nie ze stałą prędkością, ale z przyspieszeniem. Mówiąc obrazowo, depolaryzacja przyspiesza tak bardzo, że przeskakuje znak zerowy nie zauważając tego, a nawet przechodzi w polaryzację dodatnią. Po osiągnięciu szczytu (zwykle +30 mV) zaczyna się proces odwrotny – repolaryzacja, czyli przywrócenie ujemnej polaryzacji membrany.
Opiszmy krótko zjawiska elektryczne podczas przepływu potencjału czynnościowego.

Rosnąca gałąź wykresu:
- potencjał spoczynkowy - początkowy zwykły spolaryzowany stan elektroujemny błony (-70 mV);
- rosnący potencjał lokalny – depolaryzacja proporcjonalna do bodźca;
- krytyczny poziom depolaryzacji (-50 mV) - gwałtowne przyspieszenie depolaryzacji (z powodu samootwierania się kanałów sodowych), od tego punktu zaczyna się kolec - część potencjału czynnościowego o wysokiej amplitudzie;
- samowzmacniająca się, gwałtownie narastająca depolaryzacja;
- przejście znaku zerowego (0 mV) - zmiana polaryzacji membrany;
- "przeregulowanie" - polaryzacja dodatnia (inwersja lub odwrócenie ładunku membrany);
- szczyt (+30 mV) – szczyt procesu zmiany polaryzacji błony, szczyt potencjału czynnościowego.
Zstępująca gałąź wykresu:
- repolaryzacja - przywrócenie dawnej elektroujemności membrany;
- przejście znaku zerowego (0 mV) - odwrotna zmiana polaryzacji membrany na poprzednią, ujemną;
- przejście krytycznego poziomu depolaryzacji (-50 mV) - zakończenie fazy względnej ogniotrwałości (niepobudliwości) i powrót pobudliwości;
- procesy śladowe (depolaryzacja śladowa lub hiperpolaryzacja śladowa);
- przywrócenie potencjału spoczynkowego - norma (-70 mV).
Tak więc najpierw - depolaryzacja, potem - repolaryzacja. Najpierw utrata elektroujemności, potem przywrócenie elektroujemności.
2. Przepływy jonowe. W przenośni możemy powiedzieć, że naładowane jony są twórcami potencjałów elektrycznych w komórkach nerwowych. Dla wielu osób dziwnie brzmi stwierdzenie, że woda nie przewodzi prądu. Ale w rzeczywistości tak jest. Sama woda jest izolatorem, a nie przewodnikiem. W wodzie prąd elektryczny dostarczają nie elektrony, jak w metalowych drutach, ale naładowane jony: dodatnie kationy i ujemne aniony. W żywych komórkach główna „praca elektryczna” jest wykonywana przez kationy, ponieważ są one bardziej mobilne. Prądy elektryczne w komórkach to przepływy jonów.
Dlatego ważne jest, aby zdać sobie sprawę, że wszystkie prądy elektryczne, które przechodzą przez membranę, są prądami jonowymi. Po prostu nie ma prądu znanego nam z fizyki w postaci przepływu elektronów w komórkach, jak w systemach wodnych. Błędem byłoby odwoływanie się do przepływów elektronów.
Na poziomie chemicznym, opisując wzbudzenie propagujące, musimy wziąć pod uwagę, jak zmieniają się właściwości przepływającego jonu przez błonę. Najważniejsze w tym procesie jest to, że podczas depolaryzacji przepływ jonów sodu do komórki gwałtownie wzrasta, a następnie nagle zatrzymuje się na szczycie potencjału czynnościowego. Napływający sód powoduje jedynie depolaryzację, ponieważ jony sodu wnoszą ze sobą ładunki dodatnie do komórki (co zmniejsza elektroujemność). Następnie, po skoku, wypływ jonów potasu na zewnątrz znacznie się zwiększa, co powoduje repolaryzację. W końcu potas, jak wielokrotnie mówiliśmy, zabiera ze sobą ładunki dodatnie z komórki. Ładunki ujemne pozostają w większości wewnątrz ogniwa, przez co wzrasta elektroujemność. Jest to przywrócenie polaryzacji z powodu wychodzącego przepływu jonów potasu. Zauważ, że wypływ jonów potasu następuje prawie równocześnie z pojawieniem się przepływu sodu, ale rośnie powoli i trwa 10 razy dłużej. Pomimo czasu trwania przepływu potasu przez same jony, zużywa się niewiele - tylko jedna milionowa rezerwy potasu w komórce (część 0,000001).
Podsumujmy. Rosnąca gałąź wykresu potencjału czynnościowego powstaje w wyniku wejścia jonów sodu do komórki, a gałąź zstępująca wynika z wyjścia jonów potasu z komórki.
3. Kanały jonowe. Wszystkie trzy aspekty procesu wzbudzania – elektryczny, chemiczny i strukturalny – są niezbędne do zrozumienia jego istoty. Jednak wszystko zaczyna się od pracy kanałów jonowych. To stan kanałów jonowych z góry determinuje zachowanie jonów, a z kolei zachowaniu jonów towarzyszą zjawiska elektryczne. Kanały sodowe rozpoczynają proces wzbudzania.
Na poziomie struktury molekularnej otwierają się membranowe kanały sodowe. Początkowo proces ten postępuje proporcjonalnie do siły wpływu zewnętrznego, a potem staje się po prostu „niepowstrzymany” i masowy. Otwarcie kanałów umożliwia przedostanie się sodu do komórki i powoduje depolaryzację. Następnie po około 2-5 milisekundach są automatycznie zamykane. To zamknięcie kanałów nagle odcina przepływ jonów sodu do komórki, a tym samym odcina wzrost potencjału elektrycznego. Potencjalny wzrost zatrzymuje się, a na wykresie widzimy skok. To jest wierzchołek krzywej na wykresie, wtedy proces będzie przebiegał w przeciwnym kierunku. Oczywiście bardzo interesujące jest zrozumienie, że kanały sodowe mają dwie bramki i otwierają się bramką aktywacyjną i zamykają bramką dezaktywacyjną, ale należy to omówić wcześniej, w temacie „Pobudzenie”. Nie poprzestaniemy na tym.
Równolegle z otwieraniem się kanałów sodowych z niewielkim opóźnieniem następuje coraz większe otwarcie kanałów potasowych. Są powolne w porównaniu do sodu. Otwarcie dodatkowych kanałów potasowych wzmaga uwalnianie dodatnich jonów potasu z komórki. Uwalnianie potasu przeciwdziała depolaryzacji „sodowej” i powoduje przywrócenie polaryzacji (przywrócenie elektroujemności). Ale kanały sodowe wyprzedzają kanały potasowe, działają około 10 razy szybciej. W związku z tym napływ dodatnich jonów sodu do komórki wyprzedza kompensacyjny odpływ jonów potasu. A zatem depolaryzacja rozwija się szybciej niż polaryzacja przeciwstawna, spowodowana wyciekiem jonów potasu. Dlatego dopóki kanały sodowe się nie zamkną, przywrócenie polaryzacji nie rozpocznie się.
Aby przejść do zrozumienia znaczenia dynamicznego procesu wzbudzenia, tj. Aby zrozumieć jego rozkład wzdłuż błony, należy sobie wyobrazić, że opisane powyżej procesy wychwytują najpierw najbliższe, a potem wszystkie nowe, coraz bardziej odległe fragmenty błony, aż przejdą przez całą błonę. Jeśli widziałeś „żywą falę”, którą kibice na stadionie organizują wstając i kucając, to łatwo sobie wyobrazić membranową falę wzbudzenia, która powstaje dzięki sukcesywnemu przepływowi transbłonowych prądów jonowych w sąsiednich obszary.
Jako analogię lub metaforę, która może wizualnie przekazać znaczenie rozprzestrzeniającego się podniecenia, możesz wziąć obraz ognia. Rzeczywiście, rozprzestrzeniające się wzbudzenie jest jak pożar lasu, kiedy płonące drzewa pozostają na miejscu, a front ognia rozprzestrzenia się i idzie coraz dalej we wszystkich kierunkach od źródła zapłonu.
Jak będzie wyglądało zjawisko hamowania w tej metaforze?
Odpowiedź jest oczywista – hamowanie będzie wyglądało jak gaszenie pożaru, jak zmniejszenie spalania i gaszenie ognia. Ale jeśli ogień rozprzestrzenia się sam, gaszenie wymaga wysiłku. Z obszaru gaszenia sam proces gaszenia nie będzie przebiegał we wszystkich kierunkach.
Istnieją trzy możliwości gaszenia pożaru: (1) albo trzeba poczekać, aż wszystko się wypali i pożar wyczerpie wszystkie palne zapasy, (2) albo polać wodą płonące miejsca, żeby zgasły, (3) lub trzeba wcześniej podlać najbliższe obszary nietknięte przez ogień, aby się nie zapaliły.
Czy da się „ugasić” falę rozprzestrzeniającego się wzbudzenia?
Jest mało prawdopodobne, aby komórka nerwowa była w stanie „ugasić” ten „ogień” pobudzenia, które się rozpoczęło. Dlatego pierwsza metoda nadaje się tylko do sztucznej interwencji w pracę neuronów (na przykład do celów leczniczych). Okazuje się jednak, że całkiem możliwe jest „wypełnienie niektórych obszarów wodą” i zablokowanie rozprzestrzeniania się wzbudzenia.
Główną rolę w wzbudzeniu neuronu odgrywa membranowe kanały jonowe. Te kanały są dwojakiego rodzaju:
1. Niektóre (pierwszy typ) pracują nieprzerwanie i wypompowują jony sodu z neuronu i pompują jony potasu do cytoplazmy (kanały pompujące). W komórce powstaje różnica w stężeniu jonów: stężenie jonów potasu wewnątrz komórki jest około 30 razy wyższe niż stężenie jonów potasu na zewnątrz komórki (30:1). Natomiast stężenie jonów sodu wewnątrz komórki jest około 50 razy mniejsze niż stężenie jonów sodu na zewnątrz komórki (1:50). Błony każdej komórki, nie tylko nerwowej, mają tę różnicę. W rezultacie między cytoplazmą a środowiskiem zewnętrznym na błonie komórkowej powstaje potencjał: cytoplazma komórki jest naładowana ujemnie o wartość około 70 mV w stosunku do komórki zewnętrznej. Do stworzenia takiego potencjału potrzebne są tylko jony potasu (potencjał potasu).
2. Neuron, w przeciwieństwie do innych komórek, może być wzbudzany (generować potencjał czynnościowy). Główną rolę w wzbudzeniu pełni inny rodzaj kanałów jonowych (drugi typ), po otwarciu których jony sodu wpadają do komórki, a jony potasu zaczynają opuszczać komórkę przez otwarte kanały potasowe. Każdy rodzaj jonu (sodowy i potasowy) ma swój własny rodzaj kanału jonowego. Ruch jonów przez te kanały następuje wzdłuż gradientów stężeń, tj. z miejsca o wysokiej koncentracji do miejsca o niższej koncentracji.
Rozważ chemiczną podstawę powstawania i utrzymania potencjałów bioelektrycznych (potencjał spoczynkowy i potencjał czynnościowy).
Większość badaczy uważa, że zjawisko polaryzacji ogniwa elektrycznego wynika z nierównomiernego rozmieszczenia jonów K+ i Na+ po obu stronach błony komórkowej. Membrana ma selektywną przepuszczalność: wyższą dla jonów K+ i znacznie mniejszą dla jonów Na+. Ponadto w komórkach nerwowych istnieje mechanizm, który utrzymuje sód wewnątrzkomórkowy na niskim poziomie pomimo gradientu stężenia. Ten mechanizm nazywa się pompą sodową.
W spoczynku wewnętrzna strona błony komórkowej jest naładowana elektroujemnie w stosunku do powierzchni zewnętrznej. Wyjaśnia to fakt, że ilość jonów Na + wypompowanych z komórki za pomocą pompy sodowej nie jest dokładnie zrównoważona przez wejście jonów K + do komórki. W związku z tym część kationów sodu jest zatrzymywana przez wewnętrzną warstwę przeciwjonów (anionów) na zewnętrznej powierzchni błony komórkowej. Tak więc na błonach ograniczających komórki nerwowe utrzymuje się różnica potencjałów elektrycznych (transbłonowa różnica potencjałów elektrycznych); te membrany są pobudliwe elektrycznie.
W pewnych warunkach przepuszczalność membrany dla jonów Na + gwałtownie wzrasta. Po wzbudzeniu wywołanym przez ten lub inny czynnik, przepuszczalność błony komórki nerwowej (akson) selektywnie się zmienia: wzrasta selektywnie dla jonów Na + (około 500 razy) i pozostaje niezmieniona dla jonów K +. W rezultacie jony Na + wpadają do komórki. Kompensacyjny przepływ jonów K+ opuszczających komórkę jest nieco opóźniony. Powoduje to ujemny ładunek na zewnętrznej powierzchni błony komórkowej. Wewnętrzna powierzchnia membrany nabiera ładunku dodatniego; błona komórkowa zostaje ponownie naładowana (w szczególności błona aksonu, czyli włókno nerwowe) i pojawia się potencjał czynnościowy, czyli wyskok. Czas trwania kolca nie przekracza 1 ms. Ma fazę wstępującą, szczytową i opadającą. Faza zstępująca (spadek potencjału) wiąże się ze zwiększającą się przewagą uwalniania jonów K+ nad poborem jonów Na+ – potencjał błonowy powraca do normy. Po przeprowadzeniu impulsu w komórce przywracany jest stan spoczynku. W tym okresie jony Na +, które weszły do neuronu podczas wzbudzania, są zastępowane jonami K +. Przejście to zachodzi wbrew gradientowi stężeń, ponieważ w środowisku zewnętrznym otaczającym neurony jest znacznie więcej jonów Na+ niż w komórce po jej wzbudzeniu. Jak już wspomniano, przejście jonów Na+ w kierunku gradientu stężeń odbywa się za pomocą pompy sodowej, która do działania wymaga energii ATP. W końcu wszystko to prowadzi do przywrócenia początkowego stężenia kationów potasu i sodu wewnątrz komórki (aksonu), a nerw jest gotowy na przyjęcie kolejnego impulsu pobudzającego.
Potencjał spoczynkowy to różnica między ładunkami na zewnątrz i wewnątrz membrany. W spoczynku stężenie K + jest większe w komórce, a Na + - w środowisku zewnętrznym. Membrana posiada kanały Na+ i K+. Wszystkie kanały sodowe są zamknięte w spoczynku. Ponieważ na zewnątrz jest więcej sodu niż wewnątrz potasu, cytoplazma wewnątrz neuronu jest naładowana ujemnie w stosunku do płynu międzykomórkowego. Część kanałów potasowych jest otwarta, ale nie przepływa przez nie prąd, ponieważ jony potasu są zatrzymywane przez pole elektryczne powstałe w wyniku nierównomiernego rozkładu ładunku, które pozostaje stałe, dopóki przez membranę nie przepływa prąd jonów sodu . Po wzbudzeniu, w pewnym obszarze otwierają się kanały sodowe bramkowane napięciem. Sód wpada do ogniwa, ponieważ jego zewnętrzne stężenie jest większe niż wewnętrzne, a wewnętrzna strona membrany jest naładowana ujemnie, co przyciąga dodatnio naładowane jony. Ładunek wewnątrz membrany bardzo szybko staje się dodatni. Ten proces nazywa się depolaryzacją. Sód wejdzie do komórki aż do pewnej zmiany potencjału. Ze względu na duży prąd sodu wzdłuż gradientu stężeń i mały prąd potasu, membrana szybko zmienia potencjał. Następnie kanały sodowe bramkowane napięciem zamykają się i przez pewien czas nie mogą się otwierać. Ten okres nazywa się ogniotrwałością. W tym samym czasie, gdy kanały sodowe są zamknięte, dodatkowe kanały potasowe bramkowane napięciem otwierają się, wychodzący prąd potasowy gwałtownie wzrasta, a potencjał powraca nawet nieco mniej niż przed otwarciem kanałów sodowych. Kanały sodowe są przez pewien czas zamknięte i nie mogą się już otwierać. Czas ten nazywany jest czasem ogniotrwałości. W tym okresie stężenie po obu stronach błony powraca w przybliżeniu do wartości wyjściowej, dzięki dyfuzji i działaniu specjalnego enzymu - Na+/K+ATP-azy, który pompuje jony, w wyniku rozszczepiania ATP. Cały ten proces nazywa się impulsem nerwowym lub kolcem. Łatwo zrozumieć, że takie potencjalne skoki spowodują również otwarcie sąsiednich kanałów, prowadząc do falowego „ruchu” szpikulca. A okres refrakcji nie pozwoli na propagację sygnału w przeciwnym kierunku.
Potencjały czynnościowe mogą różnić się parametrami w zależności od rodzaju komórki, a nawet różnych części błony tej samej komórki. Najbardziej charakterystycznym przykładem różnic jest potencjał czynnościowy mięśnia sercowego i potencjał czynnościowy większości neuronów. Jednak następujące zjawiska leżą u podstaw każdego potencjału czynnościowego:
1. Błona żywej komórki jest spolaryzowana – jej wewnętrzna powierzchnia jest naładowana ujemnie w stosunku do zewnętrznej ze względu na to, że w roztworze przy jej zewnętrznej powierzchni znajduje się więcej dodatnio naładowanych cząstek (kationów), a przy powierzchni wewnętrznej jest więcej cząstek naładowanych ujemnie (anionów).
2. Membrana ma selektywną przepuszczalność – jej przepuszczalność dla różnych cząstek (atomów lub cząsteczek) zależy od ich wielkości, ładunku elektrycznego i właściwości chemicznych.
3. Błona komórki pobudliwej jest w stanie szybko zmienić swoją przepuszczalność dla określonego typu kationów, powodując przejście ładunku dodatniego z zewnątrz do wewnątrz.
Pierwsze dwie właściwości są charakterystyczne dla wszystkich żywych komórek. Trzeci to cecha komórek tkanek pobudliwych i powód, dla którego ich błony są zdolne do generowania i przewodzenia potencjałów czynnościowych.
Fazy potencjału działania
- Prespike to proces powolnej depolaryzacji (spadek elektroujemności komórki nerwowej) błony do krytycznego poziomu depolaryzacji (pobudzenie lokalne, odpowiedź lokalna).
- Potencjał szczytowy lub skok, składający się z części wznoszącej (depolaryzacja błony) i części opadającej (repolaryzacja błony).
- Ujemny potencjał śladowy - od krytycznego poziomu depolaryzacji do początkowego poziomu polaryzacji błony (depolaryzacja śladowa).
- Dodatni potencjał śladowy - wzrost potencjału błonowego i jego stopniowy powrót do pierwotnej wartości (hiperpolaryzacja śladowa).
Polaryzacja błony żywej komórki wynika z różnicy w składzie jonowym jej wewnętrznej i zewnętrznej strony. Gdy komórka znajduje się w stanie spokojnym (niewzbudzonym), jony po przeciwnych stronach błony tworzą względnie stabilną różnicę potencjałów, zwaną potencjałem spoczynkowym. Jeżeli do żywej komórki wprowadzi się elektrodę i zmierzy się spoczynkowy potencjał błonowy, będzie on miał wartość ujemną (rzędu -70 - -90 mV). Wyjaśnia to fakt, że całkowity ładunek po wewnętrznej stronie membrany jest znacznie mniejszy niż po zewnętrznej, chociaż obie strony zawierają zarówno kationy, jak i aniony. Na zewnątrz - o rząd wielkości więcej jonów sodu, wapnia i chloru, wewnątrz - jony potasu i ujemnie naładowane cząsteczki białek, aminokwasy, kwasy organiczne, fosforany, siarczany. Należy rozumieć, że mówimy o ładunku powierzchni membrany - ogólnie środowisko zarówno wewnątrz jak i na zewnątrz komórki jest naładowane neutralnie.
Potencjał błonowy może się zmieniać pod wpływem różnych bodźców. Sztucznym bodźcem może być prąd elektryczny przyłożony przez elektrodę do zewnętrznej lub wewnętrznej strony membrany. W warunkach naturalnych bodziec jest często sygnałem chemicznym z sąsiednich komórek, przechodzącym przez synapsę lub poprzez dyfuzyjną transmisję przez ośrodek międzykomórkowy. Przesunięcie potencjału błonowego może nastąpić w kierunku ujemnym (hiperpolaryzacja) lub dodatnim (depolaryzacja).
W tkance nerwowej potencjał czynnościowy z reguły występuje podczas depolaryzacji - jeśli depolaryzacja błony neuronowej osiągnie lub przekroczy pewien poziom progowy, komórka jest podekscytowana, a fala sygnału elektrycznego rozchodzi się z jej ciała do aksonów i dendrytów . (W rzeczywistych warunkach na ciele neuronu powstają zwykle potencjały postsynaptyczne, które bardzo różnią się od potencjału czynnościowego w naturze - na przykład nie przestrzegają zasady „wszystko albo nic”. Potencjały te są przekształcane w potencjał czynnościowy w specjalnej części membrany - wzgórek aksonu, dzięki czemu potencjał czynnościowy nie rozciąga się na dendryty).
Wynika to z faktu, że na błonie komórkowej znajdują się kanały jonowe - cząsteczki białka, które tworzą w błonie pory, przez które jony mogą przechodzić z wnętrza błony na zewnątrz i odwrotnie. Większość kanałów jest jono-specyficznych - kanał sodowy przepuszcza prawie tylko jony sodu i nie przepuszcza innych (zjawisko to nazywa się selektywnością). Błona komórkowa tkanek pobudliwych (nerwów i mięśni) zawiera dużą liczbę bramkowanych napięciem kanałów jonowych, które mogą szybko reagować na zmianę potencjału błonowego. Depolaryzacja błony powoduje przede wszystkim otwarcie kanałów sodowych bramkowanych napięciem. Gdy jednocześnie otwiera się wystarczająca ilość kanałów sodowych, dodatnio naładowane jony sodu przepływają przez nie do wnętrza membrany. Siłą napędową w tym przypadku jest gradient stężeń (na zewnątrz membrany jest dużo więcej dodatnio naładowanych jonów sodu niż wewnątrz ogniwa) oraz ładunek ujemny wewnątrz membrany). Przepływ jonów sodu powoduje jeszcze większą i bardzo szybką zmianę potencjału błonowego, który nazywamy potencjałem czynnościowym (w literaturze specjalistycznej jest to oznaczane PD).
Zgodnie z prawem „wszystko albo nic”, błona komórkowa tkanki pobudliwej albo w ogóle nie reaguje na bodziec, albo reaguje z maksymalną możliwą siłą w danej chwili. Oznacza to, że jeśli bodziec jest zbyt słaby, a próg nie zostanie osiągnięty, potencjał czynnościowy w ogóle nie powstaje; jednocześnie bodziec progowy wywoła potencjał czynnościowy o tej samej amplitudzie, co bodziec powyżej progu. Nie oznacza to, że amplituda potencjału czynnościowego jest zawsze taka sama - ten sam odcinek błony, będąc w różnych stanach, może generować potencjały czynnościowe o różnych amplitudach.
W spoczynkowym neuronie kanały sodowe błony są zamknięte (jednocześnie potencjał spoczynkowy wynosi 70 mV i ujemność w cytoplazmie). Jeśli potencjał błony jest zdepolaryzowany (zmniejsza polaryzację błony) o około 10 mV, otwiera się kanał jonów sodu. Ponadto kanał posiada rodzaj przesłony, która reaguje na potencjał membrany, otwierając ten kanał po osiągnięciu potencjału o określonej wartości (kanał zależny od potencjału).
Gdy tylko kanał się otworzy, jony sodu wpadają do cytoplazmy neuronu z ośrodka międzykomórkowego, którego jest około 50 razy więcej niż w cytoplazmie (prawem fizycznym jest ruch wzdłuż gradientu stężenia). W ten sposób jony sodu, które są naładowane dodatnio, wchodzą do neuronu. Prąd jonów sodu wchodzących przez membranę przesunie potencjał membrany w kierunku depolaryzacji, czyli zmniejszy polaryzację membrany. Potencjał w błonie wzrośnie, otwierając coraz więcej kanałów sodowych. Ale ten potencjał zatrzymuje swój wzrost, gdy tylko osiągnie +55 mV. Potencjał ten odpowiada stężeniu jonów sodu obecnych w neuronie i poza nim (potencjał równowagi sodu). (W stanie spoczynku -70 mV, amplituda bezwzględna około 125 mV).
Zatem w spoczynku ogniwo zachowuje się jak „elektroda potasowa”, a po wzbudzeniu zachowuje się „jak elektroda sodowa”. Jednak po osiągnięciu przez potencjał na błonie maksymalnej wartości +55 mV, kanał jonów sodowych od strony zwróconej do cytoplazmy jest blokowany przez specjalną cząsteczkę białka (receptor) po 0,5–1 ms i nie zależy od potencjał na membranie. Membrana staje się nieprzepuszczalna dla jonów sodu. Aby potencjał błonowy powrócił do swojego pierwotnego stanu, tj. W stanie spoczynku konieczne jest, aby prąd cząstek dodatnich, czyli jonów potasu, opuścił komórkę. Zaczynają wychodzić przez otwarte kanały potasowe. (Jony potasu gromadzą się w komórce w stanie spoczynku, dlatego po otwarciu kanałów potasowych jony te opuszczają neuron, przywracając potencjał błonowy do pierwotnego poziomu (poziom spoczynkowy). Neuron powraca do stanu spoczynkowego (-70 mV) i neuron przygotowuje się do kolejnego aktu wzbudzenia.
Po wzbudzeniu neuron przez pewien czas znajduje się w stanie absolutnej refrakcji, gdy żadne sygnały nie mogą go ponownie wzbudzić, następnie wchodzi w fazę względnej refrakcji, w której mogą go wzbudzać wyjątkowo silne sygnały (w tym przypadku amplituda AP będzie być niższy niż zwykle). Okres refrakcji występuje z powodu inaktywacji szybkiego prądu sodowego, czyli inaktywacji kanałów sodowych.