Описание презентации по отдельным слайдам:
1 слайд
Описание слайда:
2
слайд

Описание слайда:
Цели и задачи: Познакомиться с понятием рождаемость и смертность Сформировать знания о динамике численности групп организмов, о связи продолжительности жизни особей популяции Рассмотреть кривые выживания и обуславливающие их механизмы Дать представление о роли математики в современном мире, о способах применения математики в различных сферах (экология)
3
слайд

Описание слайда:
Рождаемость Рождаемость характеризует способность популяции к увеличению численности за счёт размножения особей Показатель рождаемости – это число особей (яиц семян), родившихся (вылупившихся, отложенных) в популяции за определённый промежуток времени Максимальная рождаемость – это теоретический максимум скорости образования новых особей в идеальных условиях, когда отсутствуют внешние факторы, сдерживающие процессы размножения (определяется физиологической плодовитостью и общим количеством потомства за определённый срок)
4
слайд

Описание слайда:
Экологическая рождаемость – это скорость возрастания численности популяции при фактически сложившихся условиях жизни рассматриваемой группы особей, не постоянна и изменяется в зависимости от физических условий среды и состава популяции
5
слайд

Описание слайда:
Рождаемость – это скорость размножения. Скорость зависит от времени, следовательно рождаемость есть функция времени. Функцию можно задать несколькими способами. Один из способов – графический.
6
слайд

Описание слайда:
Задача: определите, какой график соответствует длительному росту численности бактерий, помещенных на питательную среду в чашке Петри Число бактерий время Число бактерий Число бактерий А В С Рост численности бактерий в чашке Петри. А,В,С – опыты разной продолжительности
7
слайд

Описание слайда:
Для того чтобы удобнее было сравнивать рождаемость в популяциях разной численности скорость нарождения новых особей V=ΔN/ΔT (ΔN – приплод за время ΔT) относят к общей численности популяции (N). Полученную величину V/N= ΔN/(ΔT·N) называют удельной рождаемостью. Поскольку в течение промежутка времени ΔT величина рождаемости может меняться, ее стараются рассматривать на более коротких временных интервалах.
8
слайд

Описание слайда:
Различие между абсолютной (максимальной) и удельной рождаемостью в популяции рассмотрим на примерах: Пусть популяция состоящая из 50 простейших организмов в некотором объеме воды, увеличивается путем деления. Через 1 час ее численность возросла до 150 особей. Абсолютная рождаемость составила … особей в 1 час, а удельная – … особи. 100; 2 Пусть в городе с населением 10000 человек за год появилось 400 новорожденных. Абсолютная рождаемость составит … человек в год, а удельная - … . 400; 0,04
9
слайд

Описание слайда:
Смертность Смертность отражает гибель особей в популяции в данный период или число смертей в единицу времени Смертность – это показатель противоположный рождаемости, выражается чаще в виде относительной или удельной величины (процент особей, погибших в единичный отрезок времени, или их доля от начальной численности группы) При повышении смертности численность популяции сокращается и тогда стимулируются процессы, отвечающие за рождаемость. Таким образом достигается оптимальная численность популяции.
10
слайд

Описание слайда:
Изменения численности генераций при постоянной рождаемости (1000 организмов/год) и разной смертности Примечания: При расчетах принимать во внимание только целые числа 2) При начальной численности генерации 1000 особей и постоянной смертности М(%) расчет числа особей в следующем возрасте можно осуществлять по формуле 1000·(1-0,01М).
11
слайд

Описание слайда:
Каждый организм характеризуется своей индивидуальной продолжительностью жизни, которая может меняться в некоторых присущих данному виду пределах. Средняя продолжительность жизни группы организмов определяется величиной смертности. Чем больше смертность, тем меньше средняя продолжительность жизни группы, и наоборот.
12
слайд

Описание слайда:
Выживаемость Выживаемость – средняя для популяции вероятность сохранения особей каждого поколения за определённый срок. Каждый вид имеет свою свойственную ему кривую выживания. Три типа кривых выживания: выпуклая – смертность резко повышается лишь к концу жизни, а до этого она остаётся низкой. вогнутая – смертность очень высока на ранних стадиях жизни. прямая (сигмовидная) линия – коэффициент смертности остаётся постоянным на протяжении всей жизни
 Демографическая структура популяции и ее динамика Половое размножение возникало многократно, Y(Z) – хромосома не менее пяти раз. Первичное соотношение полов. Определяется генетическими механизмами. Объяснение Фишера
Демографическая структура популяции и ее динамика Половое размножение возникало многократно, Y(Z) – хромосома не менее пяти раз. Первичное соотношение полов. Определяется генетическими механизмами. Объяснение Фишера
 Гомогаметный и гетерогаметный пол (самцы (XY) у большинства, самки (WZ) у птиц, бабочек). Тритоны: у гребенчатого гомогаметны – самки, обыкновенного – самцы.
Гомогаметный и гетерогаметный пол (самцы (XY) у большинства, самки (WZ) у птиц, бабочек). Тритоны: у гребенчатого гомогаметны – самки, обыкновенного – самцы.
 Гаплодиплоидия у перепончатокрылых и других (у некоторых паутинных клещей, клопов, короедов и коловраток) 2 n 2 n n семяприёмник
Гаплодиплоидия у перепончатокрылых и других (у некоторых паутинных клещей, клопов, короедов и коловраток) 2 n 2 n n семяприёмник
 У лесного лемминга самки ХХ и ХY (с определенной Х) У копытного – самки ХХ и ХО (с определенной Х) У обыкновенной слепушонки XX имеют оба пола У горной слепушонки XO имеют оба пола
У лесного лемминга самки ХХ и ХY (с определенной Х) У копытного – самки ХХ и ХО (с определенной Х) У обыкновенной слепушонки XX имеют оба пола У горной слепушонки XO имеют оба пола
 Вторичное соотношение полов избирательность яйцеклеток, особенности среды матки, влияющие на вероятность оплодотворения, различия в частоте имплантации температурное определение пола детенышей Пример: 85% самцов у сайгаков после массового истребления последних Третичное соотношение полов Устанавливается в результате дифференцированной постнатальной смертности.
Вторичное соотношение полов избирательность яйцеклеток, особенности среды матки, влияющие на вероятность оплодотворения, различия в частоте имплантации температурное определение пола детенышей Пример: 85% самцов у сайгаков после массового истребления последних Третичное соотношение полов Устанавливается в результате дифференцированной постнатальной смертности.
 ТИПЫ ДИНАМИКИ ПОЛОВОЙ СТРУКТУРЫ Большаков и Кубанцев (1984) выделяют 4 типа динамики половой структуры. 1). Неустойчивый половой состав, соотношение полов (СП) меняется в разных местообитаниях и во времени, причем как вторичное, так и третичное СП. Характерен для животных с высокой плодовитостью и смертностью (насекомоядные). 2). Преобладание самцов. Характерны низкая плотность, забота о потомстве (хищные). 3). Преобладание самок в третичном СП. Номадные полигамы, образующие скопления (копытные, ластоногие). 4). Постоянство СП приблизительно 1: 1. Узкоспециализированных, стенобионтных видов (выхухоль, крот, бобр).
ТИПЫ ДИНАМИКИ ПОЛОВОЙ СТРУКТУРЫ Большаков и Кубанцев (1984) выделяют 4 типа динамики половой структуры. 1). Неустойчивый половой состав, соотношение полов (СП) меняется в разных местообитаниях и во времени, причем как вторичное, так и третичное СП. Характерен для животных с высокой плодовитостью и смертностью (насекомоядные). 2). Преобладание самцов. Характерны низкая плотность, забота о потомстве (хищные). 3). Преобладание самок в третичном СП. Номадные полигамы, образующие скопления (копытные, ластоногие). 4). Постоянство СП приблизительно 1: 1. Узкоспециализированных, стенобионтных видов (выхухоль, крот, бобр).
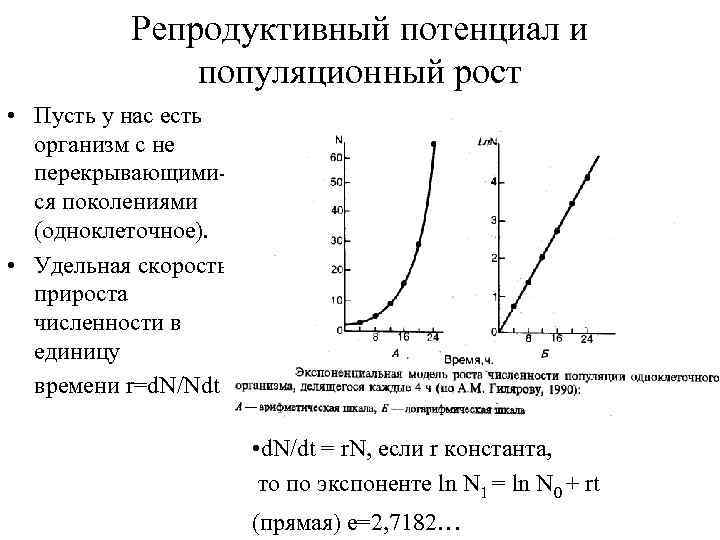 Репродуктивный потенциал и популяционный рост Пусть у нас есть организм с не перекрывающимися поколениями (одноклеточное). Удельная скорость прироста численности в единицу времени r=d. N/Ndt d. N/dt = r. N, если r константа, то по экспоненте ln N 1 = ln N 0 + rt (прямая) e=2, 7182…
Репродуктивный потенциал и популяционный рост Пусть у нас есть организм с не перекрывающимися поколениями (одноклеточное). Удельная скорость прироста численности в единицу времени r=d. N/Ndt d. N/dt = r. N, если r константа, то по экспоненте ln N 1 = ln N 0 + rt (прямая) e=2, 7182…
 А что будет, если ресурсы ограничены и скорость роста меняется в зависимости от численности? В основе логистической модели лежит линейное снижение скорости удельного роста при увеличении численности
А что будет, если ресурсы ограничены и скорость роста меняется в зависимости от численности? В основе логистической модели лежит линейное снижение скорости удельного роста при увеличении численности
 Впервые открыта бельгийским математиком Ферхюльстом (Verhulst, 1838) Переоткрыта Пирлом и Ридом (Pearl, Read, 1920)
Впервые открыта бельгийским математиком Ферхюльстом (Verhulst, 1838) Переоткрыта Пирлом и Ридом (Pearl, Read, 1920)
 Типы динамики численности Давайте обратим внимание на правую часть графика зависимости смертности и рождаемости при логистической кривой
Типы динамики численности Давайте обратим внимание на правую часть графика зависимости смертности и рождаемости при логистической кривой
 Соотношение плодовитости и смертности В правой части логистической кривой наблюдается равновесие между рождаемостью и смертностью Действительно присутствует установление соответствия между ними в эволюционных масштабах. Например, у луны-рыбы 300 миллионов пелагических икринок, а у акул - несколько яиц. Снижение плодовитости коррелирует с заботой о потомстве. У видов выкармливающих потомков плодовитость зависит от обеспеченности кормом. Плодовитость обратно пропорциональна продолжительности жизни
Соотношение плодовитости и смертности В правой части логистической кривой наблюдается равновесие между рождаемостью и смертностью Действительно присутствует установление соответствия между ними в эволюционных масштабах. Например, у луны-рыбы 300 миллионов пелагических икринок, а у акул - несколько яиц. Снижение плодовитости коррелирует с заботой о потомстве. У видов выкармливающих потомков плодовитость зависит от обеспеченности кормом. Плодовитость обратно пропорциональна продолжительности жизни
 С. А. Северцов (1941, Наумов, 1954) выделял три типа динамики численности – стабильный, лабильный и эфемерный Стабильный – виды с большой продолжительностью жизни, низкой плодовитостью, поздней половозрелостью. Период 10 -20 лет (копытные, китообразные, гоминиды, орлы). Колебания в разы Лабильный – более раннее созревание, относительно некрупные размеры (некоторые грызуны, зайцеобразные, некоторые хищные). Период – 5 -10 лет, колебания в 10 раз. Эфемерный – короткоживущие виды, большая плодовитость. Период – 3 -10 лет, колебания в 100 раз.
С. А. Северцов (1941, Наумов, 1954) выделял три типа динамики численности – стабильный, лабильный и эфемерный Стабильный – виды с большой продолжительностью жизни, низкой плодовитостью, поздней половозрелостью. Период 10 -20 лет (копытные, китообразные, гоминиды, орлы). Колебания в разы Лабильный – более раннее созревание, относительно некрупные размеры (некоторые грызуны, зайцеобразные, некоторые хищные). Период – 5 -10 лет, колебания в 10 раз. Эфемерный – короткоживущие виды, большая плодовитость. Период – 3 -10 лет, колебания в 100 раз.

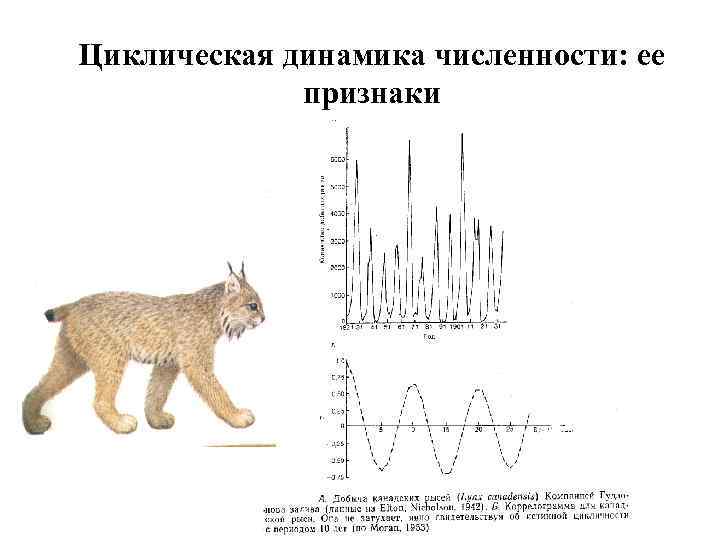

 Индекс цикличности Хенттонена Финский зоолог Henttonen с соавторами (1985) ввел индекс цикличности («амплитудности»). S - Среднее квадратичное отклонение. nлет >5 0, 16 по данным Н. В. Башениной, обычно 0, 240, 32 нецикличны, 0, 62 и выше цикличны (иногда рубеж в 0, 5) 0, 79 по данным Н. М. Окуловой 0, 85 Т. В. Кошкина, О. И. Семёнов-Тяньшанский 1, 13 для сибирского лемминга на о. Врангеля (Чернявский, Лазуткин, 2004)
Индекс цикличности Хенттонена Финский зоолог Henttonen с соавторами (1985) ввел индекс цикличности («амплитудности»). S - Среднее квадратичное отклонение. nлет >5 0, 16 по данным Н. В. Башениной, обычно 0, 240, 32 нецикличны, 0, 62 и выше цикличны (иногда рубеж в 0, 5) 0, 79 по данным Н. М. Окуловой 0, 85 Т. В. Кошкина, О. И. Семёнов-Тяньшанский 1, 13 для сибирского лемминга на о. Врангеля (Чернявский, Лазуткин, 2004)
 Факторы динамики численности 1) Факторы, независящие от плотности (экзогенные) Климатические факторы. Примеры (низкие температуры, при отсутствии снежного покрова – снижают численность и наоборот). Климат может влиять опосредованно через пищу. А) Гипотеза «климатических циклов» Чарльза Элтона (1924) В основе циклов многолетние изменения климата, погодных типов. Б) Гипотеза связи циклов с солнечной активностью. 11 летний цикл, обнаружено совпадение для непарного шелкопряда и зайцев по материалам Гудзоновой компании Предполагалось, что космическая активность действует прямо так и опосредованно. У нас эту идею развивал Анатолий Александрович Максимовым на данных по водяной полевке.
Факторы динамики численности 1) Факторы, независящие от плотности (экзогенные) Климатические факторы. Примеры (низкие температуры, при отсутствии снежного покрова – снижают численность и наоборот). Климат может влиять опосредованно через пищу. А) Гипотеза «климатических циклов» Чарльза Элтона (1924) В основе циклов многолетние изменения климата, погодных типов. Б) Гипотеза связи циклов с солнечной активностью. 11 летний цикл, обнаружено совпадение для непарного шелкопряда и зайцев по материалам Гудзоновой компании Предполагалось, что космическая активность действует прямо так и опосредованно. У нас эту идею развивал Анатолий Александрович Максимовым на данных по водяной полевке.

 Факторы, зависящие от плотности (эндогенные) Отношение потребителя и пищи Лемминговые циклы индуцируются пишей. Не только ее недостаток, но и изменения состава пищи, недостаток фосфора, калия Взаимоотношения хищник – жертва Модель Лотка-Вольтера, ее проверка Гаузе. На севере циклы есть, а на юге – нет. Анализ данных с 1871 -1949 из Норвегии показал, что в 1900 -х годах не было циклики, именно тогда велась борьба с хищниками.
Факторы, зависящие от плотности (эндогенные) Отношение потребителя и пищи Лемминговые циклы индуцируются пишей. Не только ее недостаток, но и изменения состава пищи, недостаток фосфора, калия Взаимоотношения хищник – жертва Модель Лотка-Вольтера, ее проверка Гаузе. На севере циклы есть, а на юге – нет. Анализ данных с 1871 -1949 из Норвегии показал, что в 1900 -х годах не было циклики, именно тогда велась борьба с хищниками.
 Авторегуляция 1. Информатор=Регулятор. Дрожжи –спирт. У водорослей и цианобактерий показана регуляция экзометаболитами. Химические агенты 2. Информация о плотности – поведение =регулятор. Частота контактов влияет на материнское поведение мыши. 3. Информатор – Поведение – Физиология=Регулятор. Гипотеза регуляции через стресс-реакцию Christian, 1955, 1956, 1968, Christian, Davis 1964
Авторегуляция 1. Информатор=Регулятор. Дрожжи –спирт. У водорослей и цианобактерий показана регуляция экзометаболитами. Химические агенты 2. Информация о плотности – поведение =регулятор. Частота контактов влияет на материнское поведение мыши. 3. Информатор – Поведение – Физиология=Регулятор. Гипотеза регуляции через стресс-реакцию Christian, 1955, 1956, 1968, Christian, Davis 1964
Демографическая структура популяции представляет собой численное соотношение различных категорий организмов в составе населения. Здесь главное значение имеет соотношение половых и возрастных групп, т. к.
изменение этих двух показателей существенно влияет на темпы репродукции, а значит, и на общую численность популяции и ее изменение во времени.
На демографическую структуру популяции влияют динамические показатели: рождаемость, смертность и скорость роста популяции. Большое значение имеет и продолжительность жизни отдельных особей.
Рождаемость - число особей, рождающихся в популяции за единицу времени. Смертность - это число особей, погибших в популяции в единицу времени. Однако изменения в популяции зависят не только от рождаемости и смертности, но и от скорости их иммиграции и эмиграции.
Существуют4 типа кривых, отображающих динамику популяции:
Тип "а" - S-образная кривая. Присуща популяциям, в которых высокая смертность снижает численность до уровня, когда рождаемость равна смертности и кривая выходит на "плато", что говорит об устойчивом развитии во времени;
Тип "б" - куполообразная кривая. Характерна для популяции, быстро размножившейся, а затем так же быстро погибшей в результате того, что все жизненные ресурсы оказались исчерпанными;
Тип "в" - волнообразная кривая. Наблюдается у популяций, которые быстро восстанавливают свою численность после спада, вызванного неблагоприятными факторами;
Тип "г" - кривая, демонстрирующая, как после спада численность популяции выходит на "плато", т. е. ее существование становится устойчивым.
Р. Уиттекер утверждал, что у популяции есть приспособления, благодаря которым потери популяции сокращаются, когда ее численность и ресурсы среды входят в конфликт. Он назвал этот феномен "буферностью популяции ".
Продолжительность жизни особей зависит от условий их существования. Различают физиологическую и максимальную продолжительность жизни:
- физиологическая продолжительность жизни определяется только физиологическими возможностями организма;
- максимальная продолжительность жизни - возраст, до которого может дожить лишь малая доля особей в реальных условиях среды.
По статистическим данным рождаемости и смертности получают таблицы выживания, по которым строят графики кривых выживания. Рассмотрим следующие типы выживания :
- первый тип - когда на протяжении всей жизни смертность мала, но резко возрастает в ее конце;
- второй тип - это случаи массовой гибели особей в начальный период жизни;
- третий тип - у этих популяций смертность остается постоянной в течение всей жизни.
В связи с этим выделяют типы динамики популяции :
- стабильный - характеризуется малой амплитудой и длительным периодом колебаний численности и воспринимается как стабильная система;
- лабильный - отличается закономерными колебаниями численности с периодами в 5-10 лет и более значительной амплитудой. К этому типу относятся: зайцы, некоторые хищные, птицы, рыбы, некоторые насекомые;
- эфемерный - отличается неустойчивой численностью с глубокими депрессиями, сменяющимися вспышками "массового размножения".
К механизмам устойчивости сообщества относится также и то, что популяции представлены видами с различной жизненной стратегией. Выделяют 3 фитоценотипа :
- виоленты - сильные конкуренты (например, львы);
- патиенты - особи, способные довольствоваться незначительным количеством ресурсов (верблюды);
- экстеренты - слабые конкуренты (например, шакалы).
Б.М. Миркиным (1983) была осуществлена классификация стратегий у растений, он предложил систему из 5 первичных стратегических типов :
- патиенты экотопические - переживают абиотический стресс в условиях физического неблагополучия (холод, засуха и т. д.). Взаимоотношения видов в этих условиях ослаблены;
- патиенты фитоценотические - переживают стресс под влиянием более мощных конкурентов;
- виоленты - мощные конкуренты, подавляющие жизнедеятельность других видов;
- эксплеренты типичные - кратковременно присутствуют в сообществах в период их разрушения;
- эксплеренты ложные - сезонные и флуктуационные. Вспышка их развития происходит или в определенные сезоны года, либо они реагируют на благоприятный климатический импульс.
При изучении структуры какой-либо популяции насекомых полезно, прежде всего, составить таблицу выживания для данного вида, показывающую число особей в каждой фазе жизненного цикла (см. предыдущую тему). Важную роль играет также соотношение полов. У многих насекомых число самок и самцов примерно одинаково, тогда как у множества других самки значительно превосходят самцов по численности. В ряде случаев, например у общественных насекомых, самцы составляют лишь малую долю популяции. У некоторых долгоносиков, таких, как люцерновый долгоносик и представители рода Graphognathus , самцы вообще неизвестны.
Среди других характеристик популяций следует отметить их генетическую неоднородность. В основе ее лежат различия в хромосомах, связанные с полиморфизмом определенных локусов, с перестройками хромосом и, что, возможно, важнее всего, со смешением или рекомбинацией множества сочетаний различных генов в последовательных поколениях. Генетические вариации обусловливают определенный размах морфологической (фенотипическое разнообразие) и физиологической изменчивости популяции, что исключительно важно для естественного отбора, определяющего эволюционное развитие популяции и всего вида в целом.
Регулирование численности популяций или сохранение равновесия экосистем (их кибернетика) является функцией самих экосистем и связано с взаимодействием происходящих в них процессов и составляющих их компонентов. Обычно регуляторами численности служат механизмы обратной связи, которые описываются в виде математических моделей в особом разделе экологии – системной экологии. Простой пример математической модели – это взаимодействие между плотностью популяций хищника и жертвы. При увеличении плотности популяции жертвы с некоторым запозданием растет и популяция хищника. Рост популяции жертвы делает ее более доступной для хищника, и это создает условия для постепенного роста популяции хищника. В конце концов достигается такая плотность популяции хищника, что им уничтожается значительная часть популяции жертвы. В результате популяция жертвы быстро сокращается, а это ведет к большему давлению на нее со стороны хищника, что вызывает дальнейшее уменьшение популяции жертвы. Когда она достигает очень низкого уровня, хищнику становится трудно находить жертву, и тогда популяция хищника резко уменьшается. Популяция жертвы снова получает возможность расти, и весь цикл повторяется вновь. Таким образом, численность жертвы регулируется ее естественными врагами. Вспышки численности происходят тогда, когда популяция какого-либо вида на короткое время оказывается вне влияния цикла хищник-жертва. В этих случаях популяция жертвы увеличивается быстрее, чем популяция хищника, и достигает того порога, за которым популяция хищника уже не может угнаться за нею. Происходит вспышка численности, или «эпидемия», вида-жертвы, особи которого уничтожают почти все доступные источники пищи. Ограничивающим фактором при этом служит продуктивная емкость источника корма для популяции жертвы. Как только потребности размножающейся популяции превысят эту емкость, достаточное количество пищи смогут раздобыть лишь немногие особи, и плотность популяции начнет быстро снижаться. Такого рода подъемы численности лимитируются пищевыми ресурсами.
Большую роль в жизни и адаптациях насекомых играет стабильность внешней среды. Все животные (и в особенности насекомые), населяющие местообитания с кратковременной стабильностью (например, из-за резких сезонных изменений или непредсказуемой погоды), обычно имеют небольшие размеры тела, очень подвижны, обладают высоким репродуктивным потенциалом и коротким временем генерации. Такой тип адаптации называется r -стратегией . С другой стороны, животные, в том числе и насекомые, живущие в очень стабильных условиях, чаще имеют большие размеры тела, в большей степени проявляют территориальное поведение, обладают более низким репродуктивным потенциалом и большим временем генерации. Такой тип адаптации носит название К-стратегии . Между животными, использующими одну из этих стратегий в чистом виде, находится целый спектр промежуточных форм – так называемый г-К – континуум. Можно сравнить плотность различных популяций, входящих в r-К – континуум (рис. 3), и их взаимодействие в модели хищник-жертва.
Рисунок 3 – Модели трех
различных типов роста численности и плотности популяций (по Г. Росс, Ч. Росс,
Д. Росс, 1985):
прогиб кривой в том месте, где r-отбор
и К-отбор действуют примерно в равной степени, является результатом взаимодействия
хищник-жертва
Пустынная саранча (Schistocerca gregaria ) и комнатная муха (Musca domestica ) относятся к насекомым с r-стратегией, так как условия их существования крайне непостоянны, у них мало естественных врагов, и распространяются они широко и быстро. На другом конце спектра находится яблонная плодожорка (Cydia рото nella ): она обычно обитает в стабильных условиях, развивается медленно, и плотность ее популяций довольно постоянна, т.е. это определенно организм с К-стратегией. Большинство насекомых располагается между этими крайними типами. Те из них, которые находятся ближе к типу r (например, тли), легко могут ускользать из-под регулирующего влияния естественных врагов и размножаться в катастрофических масштабах.
В сильно измененных экосистемах, например на посевах сельскохозяйственных культур, действие многих естественных регуляторов численности ограничено. В результате наблюдаются частые вспышки вредителей, уничтожающих многие культуры. Такие вспышки могут иметь катастрофические последствия, и о них известно со времен появления письменности. По мере роста численности населения земного шара и площади пахотных земель люди стали изыскивать эффективные методы борьбы с вредителями, способные заменить действие естественных регуляторов, утраченных в результате развития сельского хозяйства. В настоящее время примерно один человек из шести страдает серьезным заболеванием, переносчиками которого служат насекомые, и один из пяти недоедает в результате ущерба, наносимого насекомыми сельскохозяйственным культурам. Экологический подход к трудным проблемам восстановления сбалансированной динамики популяций насекомых, нарушенной вмешательством человека, открывает большие возможности для их решения.
| Предыдущая |
Численное соотношение различных половых и возрастных групп в составе населения рассматривается как демографическая структура популяции. Изменение этого соотношения существенно влияет на темпы репродукции и, следовательно, на общую численность популяции и ее изменение во времени.
Возрастная структура. К оценке возраста можно подходить с двух сторон. Можно оценивать абсолютный календарный возраст животных и можно определять биологический возраст, т.е. роль в популяционных процессах (продукция биомассы, участие в размножении).
Особенно отчетливо возрастные различия видны у животных с метаморфозом, включающих одну или несколько стадий. В ряде случаев разные стадии развития обитают в разных средах (амфибии или стрекозы).
При прямом развитии усложняется оценка возрастного состава, когда животные размножаются несколько раз в год, т.к. часть возрастных групп в течение года достигает половозрелости и приступает к размножению.
У видов, размножающихся один раз в год, общий спектр возрастного состава (биологический возраст) зависит от сроков наступления половозрелости. В одну группу половозрелых размножающихся особей могут входить животные разных генераций.
Знание возрастного состава является основой прогнозирования темпов роста популяции. Разным возрастным группам свойственен различный уровень смертности. Существует три варианта.
Есть виды, мало зависимые от внешних факторов смертности. У них численность разных возрастных групп (кривая выживания) находится на одном уровне, пока не достигает возраста физиологической смертности. Животных с такими особенностями не много. Это коротко живущие виды, например, поденки.
Для большинства видов характерна повышенная смертность в младших возрастных группах. Кривая выживания сначала резко падает, а потом начинает снижаться, плавно достигнув возраста относительно равномерной смертности.
При равномерном распределении смертности по возрастам характер кривой выживания имеет вид диагонально снижающейся линии. Такой вариант свойственен видам, развитие которых идет без метаморфоза при достаточной самостоятельности и устойчивости рождающегося потомства. Такая выживаемость скорее теоретическая и встречается крайне редко.
Для прогнозирования динамики численности составляются таблицы выживания, получая ожидаемую продолжительность жизни особей разных поколений.
У животных, обладающих сложным типом развития, разные стадии не только могут жить в разных средах, но и выполнять разную роль, как в жизни популяции, так и в жизни целого биоценоза, включаясь в различные трофические цепи.
У животных с прямым развитием функциональные значимые возрастные отличия не так очевидны, но также очень важны. Очевидно, что участие в репродуктивном процессе есть функция возраста и поэтому популяция, по крайней мере, делится на неполовозрелую (потенциал будущего размножения), половозрелую (осуществляющую размножение в данное время) и пострепродуктивную (закончившую свое участие в размножении) части.
Между этими частями могут быть различия в питании, что снижает внутривидовую конкуренцию,
Генерации могут отличаться по скорости роста и полового созревания. (степные пеструшки Lagurus lagurus). Быстро созревают зверьки весенних пометов, они же отличаются большей плодовитостью. Эти животные отличаются и более высоким уровнем метаболизма и меньшей продолжительностью жизни. Увеличение продолжительности жизни осенних пометов идет не за счет выживания в старости, а за счет продления физиологически юношеского периода.
Степень генетической сложности сезонных возрастов зависит от масштабов включения в размножение зверьков разных возрастов, которое отличается по годам и связано с многолетними циклами численности.
Половая структура. Соотношение полов имеет прямое отношение к интенсивности размножения и само поддержания популяции. Физиолого-экологические отличия самцов и самок также снижают внутривидовую конкуренцию. Несомненно, что поддержание биологической разно качественности послужило основой эволюционного формирования бисексуальности. Вегетативное размножение имеет задачей только наращивание биомассы и численности.
Наиболее четко половая структура выражена у членистоногих и позвоночных животных. У последних она является не только численным соотношением полов, но и различиями в биологии, связанными с формированием пространственно-этологической структуры. Половая структура динамична.
Первичное соотношение полов определяется генетическими механизмами, основывающимися на разно качественности половых хромосом (X и Y хромосомы). У млекопитающих самки имеют ХХ, а самцы ХY наборы. У птиц и бабочек наоборот. В любом случае при таком распределении первичное соотношение полов должно быть 1:1. Однако уже при оплодотворении оно может быть нарушено из-за избирательности яйцеклетки или из-за неодинаковой способности сперматозоида к оплодотворению. После оплодотворения могут быть новые влияния, например, разная частота имплантации ХХ и ХY зигот. На пол может влиять среда. Так, у рептилий на пол влияет температура инкубации. У муравьев оплодотворение происходит лишь при температуре большей 20. из них вылупляются самки. При более низких температурах откладываются неоплодотворенные яйца, из которых вылупляются только самцы. В результате среди новорожденных мы видим вторичное соотношение полов.
Третичное соотношение полов – это характеристика взрослой части популяции, складывающееся в результате дифференцированной смертности самцов и самок. Репродуктивный процесс прямо зависит именно от последнего показателя.
Выделяется четыре типа динамики половой структуры.
Неустойчивы половой состав, изменчивый в разных местообитаниях в короткие промежутки времени. Такая динамика бывает у коротко живущих животных с высокими показателями плодовитости и смертности и обширным ареалом (насекомоядные среди млекопитающих).
Преобладание самцов на фоне колеблющегося полового состава отмечено у животных с низкой плотностью и численностью. У этих животных высоко развита забота о потомстве с большими затратами энергии (хищники).
Преобладание самок в третичном соотношении полов. Самцы обычно отличаются меньшей продолжительностью жизни и в неблагоприятных условиях умирают в большем количестве. Такая половая структура свойственна Номадным полигамным млекопитающим (копытные, ластоногие). Эти животные отличаются большой продолжительностью жизни и относительно низким уровнем воспроизводства.
Наконец, для ряда групп животных характерно относительное постоянство полового состава при приблизительно одинаковом количестве самцов и самок. Такой тип свойственен узкоспециализированным стенобионтным видам, отличающимся высокой плодовитостью (выхухоль, крот, бобр)
Таким образом, реально половая структура характеризуется соотношением полов в различных возрастных группах и отражает не только интенсивность размножения, но и общий биологический потенциал популяции: ход численности, продукцию биомассы, уровень популяционных энергозатрат.
Репродуктивный потенциал и рост популяции
Скорость размножения популяции можно представить в виде удельного прироста численности за единицу времени:
где r – «мгновенная» (за короткий промежуток времени) удельная скорость роста популяции, N – ее численность, t – время изменения численности.
В этом случае неограниченный рост популяции выражается экспоненциальной кривой, описываемой уравнением
где No – исходная численность, Nt – численность в момент времени t, е (2,7182) – основание натуральных логарифмов. Если численность отложить в логарифмическом масштабе. Ее изменения будут иметь вид прямой, наклон которой определяется величиной r. Описанная экспоненциальная модель роста популяции отражает ее потенциальные возможности. Показатель r – часто называют репродуктивным потенциалом популяции или биологическим потенциалом. Описанный рост возможен только при неизменном, не зависящем от численности значении r.
Естественный рост популяции никогда таким не бывает даже в эксперименте, т.к. на него влияет целый ряд экологических условий, в том числе соотношение меняющихся значений рождаемости и смертности. Коэффициент r в таких условиях меняется. Наиболее близко естественный рост популяции описывает S-образная кривая. Уравнение логистической кривой следующее:
dN/dt=rmax N (K–N/K),
где rmax – удельная скорость роста, но в условиях исходной (минимальной) численности. По мере ее увеличения значение r падает. N –численность, K – ее предельные данные, отражающие экологическую емкость угодий. Последний параллельный оси абсцисс участок кривой означает равновесие процессов рождаемости и смертности в соответствии с данными условиями среды.