Демографическая структура популяций
Описание полового и возрастного состава популяций называют демографией (“демос” - народ, население, “графо” - пишу, описываю).
Популяции состоят из особей разного пола и возраста. Соотношение возрастных и половых групп определяет многое в общей жизнеспособности и темпах роста популяции и является важной характеристикой ее структуры.
У любой особи с возрастом закономерно изменяются характер связей со средой и устойчивость к действию отдельных факторов. У некоторых видов эти возрастные различия выражены очень резко, происходит даже смена сред обитания, характера питания, способов передвижения. Личинки стрекозы-коромысла - типично водные жители с реактивным типом движения в воде, а взрослые наземно-воздушные, с машущим полетом. Бабочки после метаморфоза переходят от грызущего типа питания к сосущему, от ползания к полету и т. п. У всех видов в ходе развития особей есть более уязвимые стадии и более устойчивые. Известно, что проростки растений, детеныши животных более чувствительны к неблагоприятным условиям, чем взрослые сформировавшиеся организмы. Самцы и самки также могут различаться по экологическим особенностям. Например, самцы кровососущих комаров вовсе не нуждаются в крови позвоночных животных, а сосут нектар цветов.
Возрастная структура популяции , т. е. соотношение в ней разных возрастных групп, зависит от двух причин: от особенностей жизненного цикла вида и от внешних условий.
Есть виды с очень простой возрастной структурой популяций, которые состоят практически из представителей одного возраста. Например, все однолетние растения весной находятся в проростках, затем примерно одновременно зацветают, дают семена и к осени отмирают. Среди животных также есть виды с однородными по возрасту популяциями, например, многие виды саранчи весной представлены личинками, ранним летом - бескрылыми неполовозрелыми особями, затем - крылатыми формами, а глубокой осенью - только яйцами, запрятанными в почве в кубышки.
Соотношение полов в популяциях определяется у некоторых видов условиями жизни или возрастом особей. У тлей , например, летом сменяют друг друга поколения, состоящие из одних самок. При неблагоприятных условиях появляются самцы. Перезимовывают оплодотворенные яйца. У ряда брюхоногих моллюсков, многощетинковых червей, рыб, ракообразных особи меняют пол с возрастом. Чаще в начале взрослой жизни животное функционирует как самец, а с увеличением размеров - как самка, но бывает и наоборот. В разных популяциях одного вида половая и возрастная структура может быть различной.
У таких видов представители разных поколений никогда не встречаются друг с другом. Численность их очень изменчива в зависимости от внешних условий. Если в уязвимый период развития наступают заморозки или засуха, происходит массовая гибель. В благоприятной же ситуации популяция может дать взрыв численности. Для видов с простой возрастной структурой изменения плотности популяции в сотни и тысячи раз - нормальное экологическое явление.
Сложная возрастная структура популяций возникает тогда, когда в ней представлены все возрастные группы, одновременно живут несколько поколений, взрослые особи размножаются многократно и имеют достаточно большую продолжительность жизни.
Возрастные пирамиды у чаек и полевок (разные популяции)
В стадах слонов или обезьян-павианов , например, есть и новорожденные, и подростки, и молодые крепнущие животные, и размножающиеся самки и самцы, и старые особи. Такие популяции не подвержены резким колебаниям численности. Критические внешние условия могут изменить их возрастной состав за счет гибели наиболее слабых, но самые устойчивые возрастные группы выживают и затем восстанавливают структуру популяции.
У зябликов в период от весеннего прилета до вылупления птенцов около 50% популяции составляют годовалые птицы, впервые начинающие размножение. На двухлетних приходится 22%, трехлетних - 12%, четырехлетних - 8%, пятилетних - 4%. На каждую тысячу птиц насчитывается только 8 особей старше 7 лет. Максимальная продолжительность жизни зяблика - 11 лет.
 Человек как биологический вид имеет сложную структуру популяций. Соотношение возрастных групп в популяциях можно наглядно выразить через пирамиду возрастов
.
Человек как биологический вид имеет сложную структуру популяций. Соотношение возрастных групп в популяциях можно наглядно выразить через пирамиду возрастов
.
Возрастные пирамиды растущей и сокращающейся популяции медоносной пчелы
Характер этой пирамиды может предсказать нам ближайшую судьбу конкретной популяции. Если в ней широкое основание, т. е. много молодых особей, узкая вершина - мало старых и достаточно представлена средняя часть, т. е. взрослые размножающиеся особи, то общая конфигурация такой пирамиды характеризует растущую популяцию. Если же основание заужено, а вершина расширена, то ждать в ближайшее время увеличения численности такой популяции не следует, в ней смертность превышает рождаемость.
В популяциях мышевидных грызунов сложность возрастной структуры закономерно возрастает от весны к осени. Весной обычно представлены перезимовавшие, еще не размножавшиеся особи. Они дают два-три приплода за лето и к осени отмирают. Особи первого приплода размножаются этим же летом и также отмирают к зиме, а их потомство и потомство от второго-третьего приплода родителей перезимовывают и размножаются только на второй год. Таким образом, популяция гораздо более разнородна в конце лета, чем весной или поздней осенью.
Естественно, что для каждого вида конфигурация возрастной пирамиды при устойчивой численности своя, поэтому нужно хорошо знать особенности его развития и взаимоотношений со средой. Например, у тех животных, у которых отсутствует забота о потомстве и в связи с этим очень высока плодовитость, основание пирамиды должно быть очень широким. Некоторые виды производят личинок в тысячи раз больше, чем их доживает до размножения. Если же забота о потомстве сильно развита, то плодовитость и доля молодых в популяциях значительно меньше.
У ряда степных и луговых злаков, таких, как ковыль, типчак или щучка , в устойчивых популяциях преобладают зрелые и старые особи. Молодые стадии растения проходят быстро и формируют мощные дерновины, которые живут очень долго, до ста лет, каждую весну давая новые побеги. Семена ежегодно попадают в почву, но прорастают только те, для которых освободится место от старых растений.
 Анализ возрастного и полового состава популяций - необходимое условие для прогноза численности тех видов, которые мы используем в дикой природе, разводим или с которыми боремся: в сельском и лесном хозяйстве, в рыбном промысле, в биологических технологиях.
Анализ возрастного и полового состава популяций - необходимое условие для прогноза численности тех видов, которые мы используем в дикой природе, разводим или с которыми боремся: в сельском и лесном хозяйстве, в рыбном промысле, в биологических технологиях.
Возрастные группы овсяницы луговой. Верхний ряд слева направо: 1 - проросток; 2, 3, 4 - молодые растения; 5, 6, 7 - взрослые растения. Нижний ряд - старые растения
У растений календарный возраст и стадия их развития (возрастное состояние) могут очень сильно не совпадать. Отдельные этапы жизненного цикла растения способны проходить с разной скоростью и, в зависимости от условий, подолгу на них задерживаться или даже проскакивать их. Если, например, о конкретном экземпляре дуба известно, что ему 30 лет, нельзя заранее сказать, что он собой представляет: уже мощное плодоносящее дерево или еще тонкий побег. Некоторые деревья в неблагоприятных условиях роста состариваются, так и не перейдя в зрелое состояние. Поэтому возрастная структура популяций у растений оценивается не по их абсолютному возрасту, а по соотношению особей на разных фазах развития. Это соотношение дает нам гораздо больше информации об условиях жизни популяции.
Например, если в уловах трески или сельди исчезают крупные взрослые рыбы и возрастной состав сдвигается в сторону молодых неполовозрелых особей, это свидетельствует о перевылове. Такой промысловой популяции надо дать отдых, чтобы молодь подросла и смогла за счет размножения пополнить популяцию. Если на лугах ценное кормовое многолетнее растение начинает встречаться только в старом возрасте, когда оно уже не способно к семенному возобновлению, следует принимать меры для омоложения состава популяции, чтобы получить в будущем устойчивый урожай этого вида.
Анализ возрастной структуры популяций человека - населения городов, поселков, деревень - крайне необходим, потому что с этим связано планирование строительства школ, детских садов, больниц, домов для престарелых и других социальных учреждений, а также расчет рабочих мест и составление общих перспектив развития данного района.
Численное соотношение различных половых и возрастных групп в составе населения рассматривается как демографическая структура популяции. Изменение этого соотношения существенно влияет на темпы репродукции и, следовательно, на общую численность популяции и ее изменение во времени.
Возрастная структура. К оценке возраста можно подходить с двух сторон. Можно оценивать абсолютный календарный возраст животных и можно определять биологический возраст, т.е. роль в популяционных процессах (продукция биомассы, участие в размножении).
Особенно отчетливо возрастные различия видны у животных с метаморфозом, включающих одну или несколько стадий. В ряде случаев разные стадии развития обитают в разных средах (амфибии или стрекозы).
При прямом развитии усложняется оценка возрастного состава, когда животные размножаются несколько раз в год, т.к. часть возрастных групп в течение года достигает половозрелости и приступает к размножению.
У видов, размножающихся один раз в год, общий спектр возрастного состава (биологический возраст) зависит от сроков наступления половозрелости. В одну группу половозрелых размножающихся особей могут входить животные разных генераций.
Знание возрастного состава является основой прогнозирования темпов роста популяции. Разным возрастным группам свойственен различный уровень смертности. Существует три варианта.
Есть виды, мало зависимые от внешних факторов смертности. У них численность разных возрастных групп (кривая выживания) находится на одном уровне, пока не достигает возраста физиологической смертности. Животных с такими особенностями не много. Это коротко живущие виды, например, поденки.
Для большинства видов характерна повышенная смертность в младших возрастных группах. Кривая выживания сначала резко падает, а потом начинает снижаться, плавно достигнув возраста относительно равномерной смертности.
При равномерном распределении смертности по возрастам характер кривой выживания имеет вид диагонально снижающейся линии. Такой вариант свойственен видам, развитие которых идет без метаморфоза при достаточной самостоятельности и устойчивости рождающегося потомства. Такая выживаемость скорее теоретическая и встречается крайне редко.
Для прогнозирования динамики численности составляются таблицы выживания, получая ожидаемую продолжительность жизни особей разных поколений.
У животных, обладающих сложным типом развития, разные стадии не только могут жить в разных средах, но и выполнять разную роль, как в жизни популяции, так и в жизни целого биоценоза, включаясь в различные трофические цепи.
У животных с прямым развитием функциональные значимые возрастные отличия не так очевидны, но также очень важны. Очевидно, что участие в репродуктивном процессе есть функция возраста и поэтому популяция, по крайней мере, делится на неполовозрелую (потенциал будущего размножения), половозрелую (осуществляющую размножение в данное время) и пострепродуктивную (закончившую свое участие в размножении) части.
Между этими частями могут быть различия в питании, что снижает внутривидовую конкуренцию,
Генерации могут отличаться по скорости роста и полового созревания. (степные пеструшки Lagurus lagurus). Быстро созревают зверьки весенних пометов, они же отличаются большей плодовитостью. Эти животные отличаются и более высоким уровнем метаболизма и меньшей продолжительностью жизни. Увеличение продолжительности жизни осенних пометов идет не за счет выживания в старости, а за счет продления физиологически юношеского периода.
Степень генетической сложности сезонных возрастов зависит от масштабов включения в размножение зверьков разных возрастов, которое отличается по годам и связано с многолетними циклами численности.
Половая структура. Соотношение полов имеет прямое отношение к интенсивности размножения и само поддержания популяции. Физиолого-экологические отличия самцов и самок также снижают внутривидовую конкуренцию. Несомненно, что поддержание биологической разно качественности послужило основой эволюционного формирования бисексуальности. Вегетативное размножение имеет задачей только наращивание биомассы и численности.
Наиболее четко половая структура выражена у членистоногих и позвоночных животных. У последних она является не только численным соотношением полов, но и различиями в биологии, связанными с формированием пространственно-этологической структуры. Половая структура динамична.
Первичное соотношение полов определяется генетическими механизмами, основывающимися на разно качественности половых хромосом (X и Y хромосомы). У млекопитающих самки имеют ХХ, а самцы ХY наборы. У птиц и бабочек наоборот. В любом случае при таком распределении первичное соотношение полов должно быть 1:1. Однако уже при оплодотворении оно может быть нарушено из-за избирательности яйцеклетки или из-за неодинаковой способности сперматозоида к оплодотворению. После оплодотворения могут быть новые влияния, например, разная частота имплантации ХХ и ХY зигот. На пол может влиять среда. Так, у рептилий на пол влияет температура инкубации. У муравьев оплодотворение происходит лишь при температуре большей 20. из них вылупляются самки. При более низких температурах откладываются неоплодотворенные яйца, из которых вылупляются только самцы. В результате среди новорожденных мы видим вторичное соотношение полов.
Третичное соотношение полов – это характеристика взрослой части популяции, складывающееся в результате дифференцированной смертности самцов и самок. Репродуктивный процесс прямо зависит именно от последнего показателя.
Выделяется четыре типа динамики половой структуры.
Неустойчивы половой состав, изменчивый в разных местообитаниях в короткие промежутки времени. Такая динамика бывает у коротко живущих животных с высокими показателями плодовитости и смертности и обширным ареалом (насекомоядные среди млекопитающих).
Преобладание самцов на фоне колеблющегося полового состава отмечено у животных с низкой плотностью и численностью. У этих животных высоко развита забота о потомстве с большими затратами энергии (хищники).
Преобладание самок в третичном соотношении полов. Самцы обычно отличаются меньшей продолжительностью жизни и в неблагоприятных условиях умирают в большем количестве. Такая половая структура свойственна Номадным полигамным млекопитающим (копытные, ластоногие). Эти животные отличаются большой продолжительностью жизни и относительно низким уровнем воспроизводства.
Наконец, для ряда групп животных характерно относительное постоянство полового состава при приблизительно одинаковом количестве самцов и самок. Такой тип свойственен узкоспециализированным стенобионтным видам, отличающимся высокой плодовитостью (выхухоль, крот, бобр)
Таким образом, реально половая структура характеризуется соотношением полов в различных возрастных группах и отражает не только интенсивность размножения, но и общий биологический потенциал популяции: ход численности, продукцию биомассы, уровень популяционных энергозатрат.
Репродуктивный потенциал и рост популяции
Скорость размножения популяции можно представить в виде удельного прироста численности за единицу времени:
где r – «мгновенная» (за короткий промежуток времени) удельная скорость роста популяции, N – ее численность, t – время изменения численности.
В этом случае неограниченный рост популяции выражается экспоненциальной кривой, описываемой уравнением
где No – исходная численность, Nt – численность в момент времени t, е (2,7182) – основание натуральных логарифмов. Если численность отложить в логарифмическом масштабе. Ее изменения будут иметь вид прямой, наклон которой определяется величиной r. Описанная экспоненциальная модель роста популяции отражает ее потенциальные возможности. Показатель r – часто называют репродуктивным потенциалом популяции или биологическим потенциалом. Описанный рост возможен только при неизменном, не зависящем от численности значении r.
Естественный рост популяции никогда таким не бывает даже в эксперименте, т.к. на него влияет целый ряд экологических условий, в том числе соотношение меняющихся значений рождаемости и смертности. Коэффициент r в таких условиях меняется. Наиболее близко естественный рост популяции описывает S-образная кривая. Уравнение логистической кривой следующее:
dN/dt=rmax N (K–N/K),
где rmax – удельная скорость роста, но в условиях исходной (минимальной) численности. По мере ее увеличения значение r падает. N –численность, K – ее предельные данные, отражающие экологическую емкость угодий. Последний параллельный оси абсцисс участок кривой означает равновесие процессов рождаемости и смертности в соответствии с данными условиями среды.
 Демографическая структура популяции и ее динамика Половое размножение возникало многократно, Y(Z) – хромосома не менее пяти раз. Первичное соотношение полов. Определяется генетическими механизмами. Объяснение Фишера
Демографическая структура популяции и ее динамика Половое размножение возникало многократно, Y(Z) – хромосома не менее пяти раз. Первичное соотношение полов. Определяется генетическими механизмами. Объяснение Фишера
 Гомогаметный и гетерогаметный пол (самцы (XY) у большинства, самки (WZ) у птиц, бабочек). Тритоны: у гребенчатого гомогаметны – самки, обыкновенного – самцы.
Гомогаметный и гетерогаметный пол (самцы (XY) у большинства, самки (WZ) у птиц, бабочек). Тритоны: у гребенчатого гомогаметны – самки, обыкновенного – самцы.
 Гаплодиплоидия у перепончатокрылых и других (у некоторых паутинных клещей, клопов, короедов и коловраток) 2 n 2 n n семяприёмник
Гаплодиплоидия у перепончатокрылых и других (у некоторых паутинных клещей, клопов, короедов и коловраток) 2 n 2 n n семяприёмник
 У лесного лемминга самки ХХ и ХY (с определенной Х) У копытного – самки ХХ и ХО (с определенной Х) У обыкновенной слепушонки XX имеют оба пола У горной слепушонки XO имеют оба пола
У лесного лемминга самки ХХ и ХY (с определенной Х) У копытного – самки ХХ и ХО (с определенной Х) У обыкновенной слепушонки XX имеют оба пола У горной слепушонки XO имеют оба пола
 Вторичное соотношение полов избирательность яйцеклеток, особенности среды матки, влияющие на вероятность оплодотворения, различия в частоте имплантации температурное определение пола детенышей Пример: 85% самцов у сайгаков после массового истребления последних Третичное соотношение полов Устанавливается в результате дифференцированной постнатальной смертности.
Вторичное соотношение полов избирательность яйцеклеток, особенности среды матки, влияющие на вероятность оплодотворения, различия в частоте имплантации температурное определение пола детенышей Пример: 85% самцов у сайгаков после массового истребления последних Третичное соотношение полов Устанавливается в результате дифференцированной постнатальной смертности.
 ТИПЫ ДИНАМИКИ ПОЛОВОЙ СТРУКТУРЫ Большаков и Кубанцев (1984) выделяют 4 типа динамики половой структуры. 1). Неустойчивый половой состав, соотношение полов (СП) меняется в разных местообитаниях и во времени, причем как вторичное, так и третичное СП. Характерен для животных с высокой плодовитостью и смертностью (насекомоядные). 2). Преобладание самцов. Характерны низкая плотность, забота о потомстве (хищные). 3). Преобладание самок в третичном СП. Номадные полигамы, образующие скопления (копытные, ластоногие). 4). Постоянство СП приблизительно 1: 1. Узкоспециализированных, стенобионтных видов (выхухоль, крот, бобр).
ТИПЫ ДИНАМИКИ ПОЛОВОЙ СТРУКТУРЫ Большаков и Кубанцев (1984) выделяют 4 типа динамики половой структуры. 1). Неустойчивый половой состав, соотношение полов (СП) меняется в разных местообитаниях и во времени, причем как вторичное, так и третичное СП. Характерен для животных с высокой плодовитостью и смертностью (насекомоядные). 2). Преобладание самцов. Характерны низкая плотность, забота о потомстве (хищные). 3). Преобладание самок в третичном СП. Номадные полигамы, образующие скопления (копытные, ластоногие). 4). Постоянство СП приблизительно 1: 1. Узкоспециализированных, стенобионтных видов (выхухоль, крот, бобр).
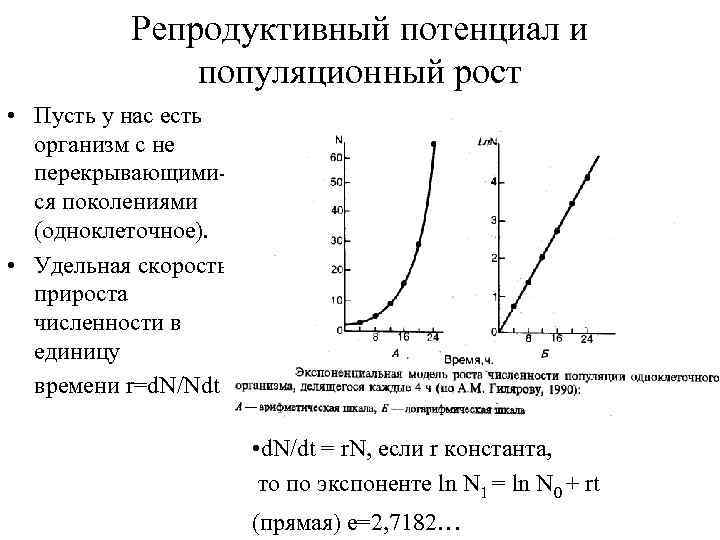 Репродуктивный потенциал и популяционный рост Пусть у нас есть организм с не перекрывающимися поколениями (одноклеточное). Удельная скорость прироста численности в единицу времени r=d. N/Ndt d. N/dt = r. N, если r константа, то по экспоненте ln N 1 = ln N 0 + rt (прямая) e=2, 7182…
Репродуктивный потенциал и популяционный рост Пусть у нас есть организм с не перекрывающимися поколениями (одноклеточное). Удельная скорость прироста численности в единицу времени r=d. N/Ndt d. N/dt = r. N, если r константа, то по экспоненте ln N 1 = ln N 0 + rt (прямая) e=2, 7182…
 А что будет, если ресурсы ограничены и скорость роста меняется в зависимости от численности? В основе логистической модели лежит линейное снижение скорости удельного роста при увеличении численности
А что будет, если ресурсы ограничены и скорость роста меняется в зависимости от численности? В основе логистической модели лежит линейное снижение скорости удельного роста при увеличении численности
 Впервые открыта бельгийским математиком Ферхюльстом (Verhulst, 1838) Переоткрыта Пирлом и Ридом (Pearl, Read, 1920)
Впервые открыта бельгийским математиком Ферхюльстом (Verhulst, 1838) Переоткрыта Пирлом и Ридом (Pearl, Read, 1920)
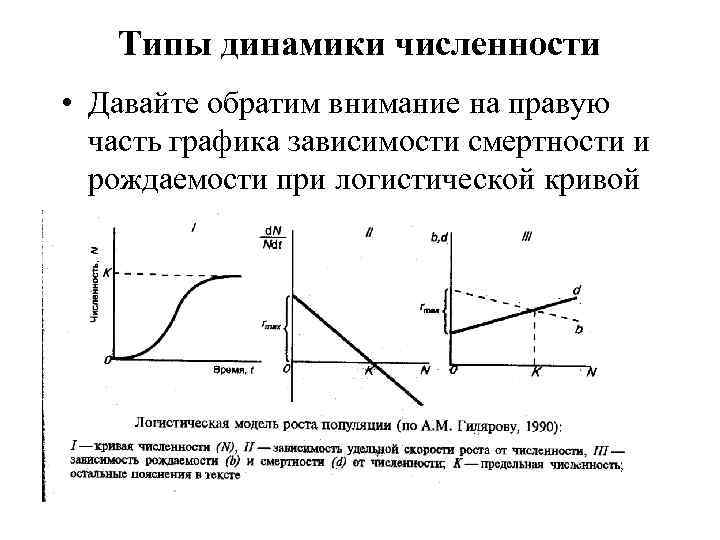 Типы динамики численности Давайте обратим внимание на правую часть графика зависимости смертности и рождаемости при логистической кривой
Типы динамики численности Давайте обратим внимание на правую часть графика зависимости смертности и рождаемости при логистической кривой
 Соотношение плодовитости и смертности В правой части логистической кривой наблюдается равновесие между рождаемостью и смертностью Действительно присутствует установление соответствия между ними в эволюционных масштабах. Например, у луны-рыбы 300 миллионов пелагических икринок, а у акул - несколько яиц. Снижение плодовитости коррелирует с заботой о потомстве. У видов выкармливающих потомков плодовитость зависит от обеспеченности кормом. Плодовитость обратно пропорциональна продолжительности жизни
Соотношение плодовитости и смертности В правой части логистической кривой наблюдается равновесие между рождаемостью и смертностью Действительно присутствует установление соответствия между ними в эволюционных масштабах. Например, у луны-рыбы 300 миллионов пелагических икринок, а у акул - несколько яиц. Снижение плодовитости коррелирует с заботой о потомстве. У видов выкармливающих потомков плодовитость зависит от обеспеченности кормом. Плодовитость обратно пропорциональна продолжительности жизни
 С. А. Северцов (1941, Наумов, 1954) выделял три типа динамики численности – стабильный, лабильный и эфемерный Стабильный – виды с большой продолжительностью жизни, низкой плодовитостью, поздней половозрелостью. Период 10 -20 лет (копытные, китообразные, гоминиды, орлы). Колебания в разы Лабильный – более раннее созревание, относительно некрупные размеры (некоторые грызуны, зайцеобразные, некоторые хищные). Период – 5 -10 лет, колебания в 10 раз. Эфемерный – короткоживущие виды, большая плодовитость. Период – 3 -10 лет, колебания в 100 раз.
С. А. Северцов (1941, Наумов, 1954) выделял три типа динамики численности – стабильный, лабильный и эфемерный Стабильный – виды с большой продолжительностью жизни, низкой плодовитостью, поздней половозрелостью. Период 10 -20 лет (копытные, китообразные, гоминиды, орлы). Колебания в разы Лабильный – более раннее созревание, относительно некрупные размеры (некоторые грызуны, зайцеобразные, некоторые хищные). Период – 5 -10 лет, колебания в 10 раз. Эфемерный – короткоживущие виды, большая плодовитость. Период – 3 -10 лет, колебания в 100 раз.
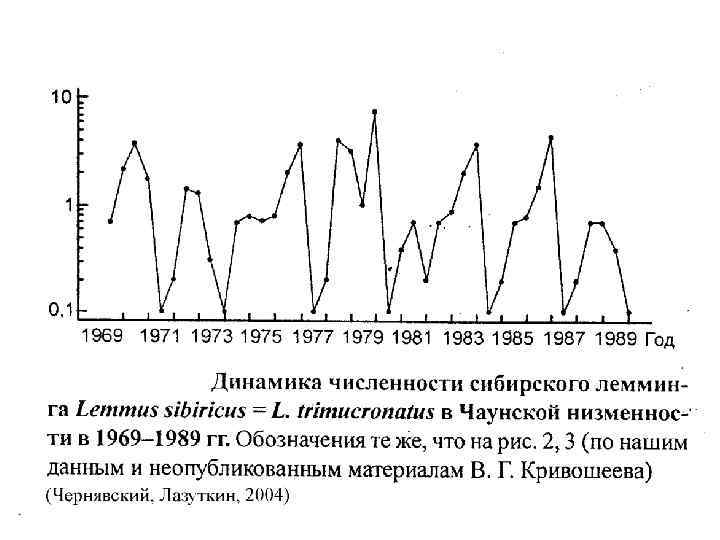


 Индекс цикличности Хенттонена Финский зоолог Henttonen с соавторами (1985) ввел индекс цикличности («амплитудности»). S - Среднее квадратичное отклонение. nлет >5 0, 16 по данным Н. В. Башениной, обычно 0, 240, 32 нецикличны, 0, 62 и выше цикличны (иногда рубеж в 0, 5) 0, 79 по данным Н. М. Окуловой 0, 85 Т. В. Кошкина, О. И. Семёнов-Тяньшанский 1, 13 для сибирского лемминга на о. Врангеля (Чернявский, Лазуткин, 2004)
Индекс цикличности Хенттонена Финский зоолог Henttonen с соавторами (1985) ввел индекс цикличности («амплитудности»). S - Среднее квадратичное отклонение. nлет >5 0, 16 по данным Н. В. Башениной, обычно 0, 240, 32 нецикличны, 0, 62 и выше цикличны (иногда рубеж в 0, 5) 0, 79 по данным Н. М. Окуловой 0, 85 Т. В. Кошкина, О. И. Семёнов-Тяньшанский 1, 13 для сибирского лемминга на о. Врангеля (Чернявский, Лазуткин, 2004)
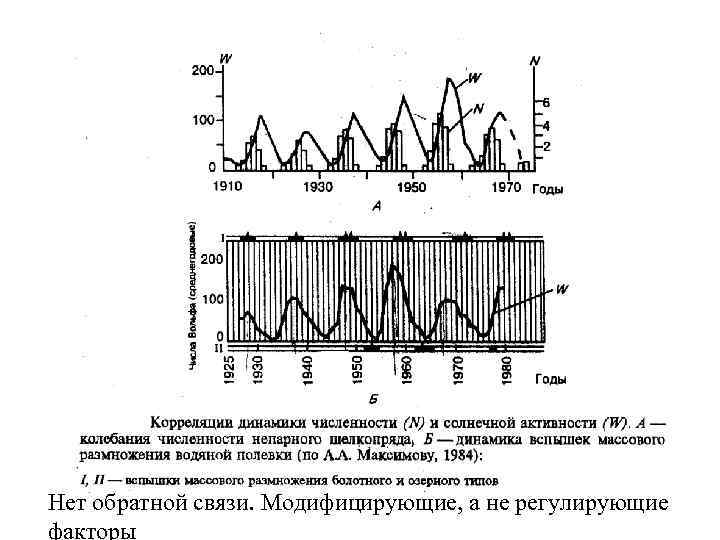
 Факторы динамики численности 1) Факторы, независящие от плотности (экзогенные) Климатические факторы. Примеры (низкие температуры, при отсутствии снежного покрова – снижают численность и наоборот). Климат может влиять опосредованно через пищу. А) Гипотеза «климатических циклов» Чарльза Элтона (1924) В основе циклов многолетние изменения климата, погодных типов. Б) Гипотеза связи циклов с солнечной активностью. 11 летний цикл, обнаружено совпадение для непарного шелкопряда и зайцев по материалам Гудзоновой компании Предполагалось, что космическая активность действует прямо так и опосредованно. У нас эту идею развивал Анатолий Александрович Максимовым на данных по водяной полевке.
Факторы динамики численности 1) Факторы, независящие от плотности (экзогенные) Климатические факторы. Примеры (низкие температуры, при отсутствии снежного покрова – снижают численность и наоборот). Климат может влиять опосредованно через пищу. А) Гипотеза «климатических циклов» Чарльза Элтона (1924) В основе циклов многолетние изменения климата, погодных типов. Б) Гипотеза связи циклов с солнечной активностью. 11 летний цикл, обнаружено совпадение для непарного шелкопряда и зайцев по материалам Гудзоновой компании Предполагалось, что космическая активность действует прямо так и опосредованно. У нас эту идею развивал Анатолий Александрович Максимовым на данных по водяной полевке.
 Факторы, зависящие от плотности (эндогенные) Отношение потребителя и пищи Лемминговые циклы индуцируются пишей. Не только ее недостаток, но и изменения состава пищи, недостаток фосфора, калия Взаимоотношения хищник – жертва Модель Лотка-Вольтера, ее проверка Гаузе. На севере циклы есть, а на юге – нет. Анализ данных с 1871 -1949 из Норвегии показал, что в 1900 -х годах не было циклики, именно тогда велась борьба с хищниками.
Факторы, зависящие от плотности (эндогенные) Отношение потребителя и пищи Лемминговые циклы индуцируются пишей. Не только ее недостаток, но и изменения состава пищи, недостаток фосфора, калия Взаимоотношения хищник – жертва Модель Лотка-Вольтера, ее проверка Гаузе. На севере циклы есть, а на юге – нет. Анализ данных с 1871 -1949 из Норвегии показал, что в 1900 -х годах не было циклики, именно тогда велась борьба с хищниками.
 Авторегуляция 1. Информатор=Регулятор. Дрожжи –спирт. У водорослей и цианобактерий показана регуляция экзометаболитами. Химические агенты 2. Информация о плотности – поведение =регулятор. Частота контактов влияет на материнское поведение мыши. 3. Информатор – Поведение – Физиология=Регулятор. Гипотеза регуляции через стресс-реакцию Christian, 1955, 1956, 1968, Christian, Davis 1964
Авторегуляция 1. Информатор=Регулятор. Дрожжи –спирт. У водорослей и цианобактерий показана регуляция экзометаболитами. Химические агенты 2. Информация о плотности – поведение =регулятор. Частота контактов влияет на материнское поведение мыши. 3. Информатор – Поведение – Физиология=Регулятор. Гипотеза регуляции через стресс-реакцию Christian, 1955, 1956, 1968, Christian, Davis 1964
К параметрам демографической структуры популяции прежде всего принадлежит соотношение полов (половой состав). Количественное соотношение полов, или половую структуру популяции, принято обозначать латинской буквой г, от термина «ratio» или, точнее сказать, «sex ratio».
Соотношение полов прежде всего зависит от биологии вида и сильно разнится у моногамов и полигамов (рис. 50, 51). Для первых типично в качестве нормы r=1: 1 с некоторыми отклонениями в ту или иную сторону в зависимости от привходящих обстоятельств. Соотношение полов у полигамов характеризуется преобладанием самок, причем эта диспропорция может быть резко выраженной. Вообще для судеб популяций и тех и других видов чрезвычайно важна значительная изменчивость коэффициента r.
Рис. 50. Пара черных журавлей - пример моногамии.
Рис. 51. Гаремы морских котиков - пример полигамии.
Среди моногамов едва ли не постоянно имеются «резервные» самцы. Они уже достигли половой зрелости, но не размножаются, а составляют репродуктивный резерв популяции. Он может сыграть очень важную роль в случае гибели взрослых размножающихся особей, тем более что смертность среди самцов бывает выше, чем среди самок. Происходящий в результате этого неравномерный урон, таящий в себе опасность нарушения нормального воспроизводства популяции, как раз и компенсируется этими резервными животными. Они занимают освободившиеся места партнеров и выравнивают процесс размножения.
Неравномерность гибели разных полов, неодинаковая их выживаемость широко распространена среди животных. Она проявляется уже на ранних стадиях эмбрионального развития, продолжается в постнатальном онтогенезе вплоть до зрелого возраста. Неизменно самки оказываются более жизнеспособными, чем самцы. В основе данного явления, вероятно, лежат какие-то физиологические и биохимические различия между особями мужского и женского пола. В молодом и зрелом возрасте большое значение приобретают отличия поведения. Самцы составляют значительно более подвижную часть популяции, они менее привязаны к убежищам, к выводкам, чаще оказываются вне убежищ и поэтому скорее становятся жертвами, хищников и непогоды. Вместе с тем самцы по тем же самым причинам в большем числе, чем самки, попадаются в капканы и ловушки. Поэтому, кстати, по результатам таких отловов трудно составить правильное представление об истинном соотношении полов в популяции, например, пушных зверей.
Выживаемость самок особенно возрастает при неблагоприятных условиях, когда популяция находится в депрессии. В результате процент женских особей далеко превышает норму. Это явление имеет очень важное адаптивное значение, поскольку от самок в первую очередь зависит восстановление подорванной численности данной популяции.
Второй параметр демографической структуры популяции - ее возрастной состав. Здесь речь идет о наличии поколений и генераций. Возрастная структура играет очень важную роль в жизни популяций. У многих видов она отличается большой сложностью. Например, в популяциях растений различаются по крайней мере четыре возрастные фазы, или группы, соответствующие известным периодам онтогенеза:
1. Латентный период - фаза первичного покоя. В эту группу входят семена, плоды и другие зачатки, которые еще не тронулись в рост и могут годами находиться в состоянии первичного покоя. Такого рода зачатки составляют очень важный семенной резервный фонд для этих растений, реализуемый при возникновении оптимальных условий произрастания.
2. Виргинильный (девственный или юношеский) период - охватывает группу растительных организмов в период от прорастания зачатков до образования генеративных органов. Период включает три стадии: всходы, ювенильную (юношескую), имматурную (переходную от ювенильного к взрослому, генеративному состоянию).
3. Генеративный период - фаза размножения растений семенами или другими разносимыми зачатками, т. е. время полового размножения.
4. Сенильный, или старческий, период - к нему принадлежат особи, закончившие половое размножение и способные только вегетировать.
Для продолжения рода у растений чрезвычайно важна их семенная продуктивность, урожайность. Количество семян, продуцируемых одним растением, может быть очень большим, прежде всего у видов с мелкими семенами. Например, некоторые представители семейства орхидных приносят до 50 тыс. семян. Чрезвычайно большой плодовитостью отличаются крупные деревья. На отдельных изолированно растущих елях родится до 100 шишек и более (т. е. 0,5 кг семян и свыше того), на соснах - до 7 тыс. шишек, на сибирских кедрах-1-3 тыс. шишек (15-45 кг «орешков»). На ясене однажды насчитали 224 тыс. семян. Береза бородавчатая приносит до 200 тыс. сережек или 80 млн семян (до 13,5 кг). В насаждениях плодовитость каждого отдельного дерева, конечно, значительно меньше, но их суммарная производительность на единицу площади очень велика, особенно в наиболее урожайные годы. Эти максимальные показатели отражены в таблице 3.
Страницы: 1Описание полового и возрастного состава популяций называют демографией (“демос” – народ, население, “графо” – пишу, описываю).
Под демографической структурой популяции понимают прежде всего ее половой и возрастной состав. Кроме того, принято говорить о пространственной структуре популяции - то есть об особенностях размещения особей популяции в пространстве.
Знание структуры популяции позволяет исследователю сделать выводы о ее благополучии или неблагополучии. Например, если в популяции отсутствуют генеративные (то есть способные дать потомство) особи и при этом много старовозрастных (сенильных) особей, то можно сделать неблагоприятный прогноз. У такой популяции может не быть будущего. Структуру популяции желательно изучать в динамике: зная ее изменение в течение нескольких лет, можно намного более уверенно говорить о тех или иных тенденциях.
Возрастная структура популяции . Этот тип структуры связан с соотношением особей различных возврастов в популяции. Особи одного возраста принято объединять в когорты , то есть возрастные группы.
Возрастная структура популяций растений описана очень подробно. В ней выделяют (по Т.А. Роботнову) следующие возрасты (возрастные группы организмов):
латентный период - состояние семени;
прегенеративный период (включает состояния проростка, ювенильного растения, имматурного растения и виргинильного растения);
генеративный период (обычно подразделяется на три подпериода - молодых, зрелых и старых генеративных особей);
постгенеративный период (включает состояния субсенильного растения, сенильного растения и фазу отмирания).
Разумеется, при этом возникает проблема соотношения календарного и биологического возраста. Принадлежность к определенному возрастному состоянию определяется по степени выраженности определенных морфологических (например, степень расчлененности сложного листа) и физиологических (например, способность дать потомство) признаков. Таким образом фиксируется, прежде всего, биологический возраст особи. Биологический возраст имеет для эколога большее значение, так как именно он определяет роль особи в популяционных процессах. В то же время, как правило, существует взаимосвязь между биологическим и календарным возрастом.
В популяциях животных также можно выделить различные возрастные стадии. Например, насекомые, развивающиеся с полным метаморфозом, проходят стадии яйца, личинки, куколки, имаго (взрослого насекомого). У других животных (развивающихся без метаморфоза) также можно выделить различные возрастные состояния, хотя границы между ними могут быть и не настолько четкими.
Характер возрастной структуры (или как говорят, возрастного спектра) популяции зависит от типа так называемой кривой выживания, свойственной данной популяции. Кривая выживания отражает уровень смертности в различных возрастных группах. Так, если уровень смертности не зависит от возраста особей, то кривая выживания представляет собой снижающуюся линию (см. рисунок, тип I). То есть отмирание особей происходит в данном типе равномерно, коэффициент смертности остается постоянным на протяжении всей жизни. Такая кривая выживания свойственна видам, развитие которых происходит без метаморфоза при достаточной устойчивости рождающегося потомства. Этот тип принято называть типом гидры - для нее свойственна кривая выживания, приближающаяся к прямой линии.
У видов, для которых роль внешних факторов в смертности невелика, кривая выживания характеризуется небольшим понижением до определенного возраста, после которого происходит резкое падение в следствие естественной (физиологический) смертности. Тип II на рисунке. Близкий к этому типу характер кривой выживания свойственен человеку (хотя кривая выживания человека несколько более пологая и, таким образом, является чем-то средним между типами I и II). Этот тип носит названия типа дрозофиллы: именно его демонстрируют дрозофиллы в лабораторных условиях (не поедаемые хищниками).
Для очень многих видов характерна высокая смертность на ранних стадиях онтогенеза. У таких видов кривая выживания характеризуется резким падением в области младших возрастов. Особи, пережившие "критический" возраст, демонстрируют низкую смертность и доживают до больших возрастов. Тип носит название типа устрицы. Тип III на рисунке.
Изучение кривых выживания представляет большой интерес для эколога. Оно позволяет судить о том, в каком возрасте тот или иной вид наиболее уязвим. Если действие причин, способных изменить рождаемость или смертность, приходится на наиболее уязвимую стадию, то их влияние на последующее развитие популяции будет наибольшим. Эту закономерность необходимо учитывать при организации охоты или в борьбе с вредителями.
Половая структура популяции .
О половой структуре популяции можно говорить, разумеется, только если речь идет о раздельнополом (бисексуальном) виде. Бисексуальность играет огромную роль в поддержании генетической разнокачественности особей популяции. Значение генетической разнокачественности для устойчивости популяции будет подробно раскрыто в следующем уроке. Сейчас же отметим, что половая структура, то есть соотношение полов, имеет прямое отношение к воспроизводству популяции и ее устойчивости.
Принято выделять первичное, вторичное и третичное соотношение полов в популяции. Первичное соотношение полов определяется генетическими механизмами - равномерностью расхождения половых хромосом. Например, у человека XY-хромосомы определяют развитие мужского пола, а XX - женского. В этом случае первичное соотношение полов 1:1, то есть равновероятно.
Вторичное соотношение полов - это соотношение полов на момент рождения (среди новорожденных). Оно может существенно отличаться от первичного по целому ряду причин: избирательность яйцеклеток к сперматозоидам, несущим X- или Y-хромосому, неодинаковой способностью таких сперматозоидов к оплодотворению, различными внешними факторами. Например, зоологами описано влияние температуры на вторичное соотношение полов у рептилий. Аналогичная закономерность характерна и для некоторых насекомых. Так, у муравьев оплодотворение обеспечивается при температуре выше 20 С, а при более низких температурах откладываются неоплодотворенные яйца. Из последних вылупляются самцы, а из оплодотворенных - преимущественно, самки.
Третичное соотношение полов - это соотношение полов среди взрослых животных.